
Abstract
Seabreams from two genera, Spondyliosoma and Spicara, have a unique life-history that combines nesting, paternal care, and protogyny. Whereas protogyny is a common sexual system among seabreams, all, except Spondyliosoma and Spicara spp., are broadcast spawners. We describe the nesting behaviour of Spondyliosoma emarginatum as a basis for analysing this unusual life-history combination. We remotely filmed males on a nesting site with over 80 nests in South Africa. It has been postulated, based on morphometric data and general observations, that males invest heavily in nest-building, courtship, and egg-guarding, although these behaviours had never been documented. Energy intensive behaviours, including the clearing of invertebrate invaders (7 counts h−1) and fanning of nests (86 counts h−1), remained constant before and after the appearance of eggs. Males spent 79.2% of their time on the nest. The remainder was spent chasing other fish species and neighbouring males (12 counts h−1) and avoiding predation. Paired spawning events proceeded in batches. Nest take-overs and sneaker male attempts were recorded. The young leave the nest immediately after hatching. Males did not feed during the 67-day nesting period. Energetic costs of nest maintenance, starvation, and predation risk explain the condensed (relative to other seabreams) 51-day spawning season. S. emarginatum meets the requirements of the SAM as nest defence requires large male body size. Their short lifespan classifies Spondyliosoma spp. as opportunists, which may explain their success across a wide range of biogeographic zones from the cold temperate North Atlantic to the subtropical western Indian Ocean. These characteristics set Spondyliosoma spp. apart from the bulk of the seabreams, which are long-lived, broadcast-spawning, periodic strategists.
Similar content being viewed by others


Introduction
Hermaphroditism, nest-building, and paternal care are phylogenetically versatile traits that add to the remarkable diversity of life-history strategies among fishes. The co-occurrence of these particular traits, however, is rare, partly because of their vastly unequal frequencies between fishes of freshwater and marine environments. Functional hermaphroditism (an individual acts as both a male and a female at some stage in its life) is very rare among freshwater fishes compared to saltwater fishes (De Mitcheson and Liu 2008; Kuwamura et al. 2020; Pla et al. 2021), but nest-building and parental care are considerably more frequent among freshwater than saltwater fishes (Blumer 1982).
Sequential hermaphroditism evolved independently several times in fish. Variations exist between estimates on the total number of families containing hermaphroditic species due to difficulties in diagnosing hermaphroditism in species (Pla et al. 2021). One estimate is that it is found among fishes from 41 families, mostly on tropical continental shelves (Kuwamura et al. 2020). Even within a single family, the Sparidae, hermaphroditism evolved a few times independently (Pla et al. 2020).
Parental care was defined as “any investment by the parent in an individual offspring that increases the offspring’s chance of surviving and hence reproductive success at the cost of the parent’s ability to invest in other offspring” (Trivers 1972). Models for the evolution of parental care were built on the investment hypothesis, which draws on two important concepts: there are costs associated with parental care (Trivers 1972), and these costs differ between the sexes (Bateman 1948). Whereas females invest heavily in oocytes, males spend more energy on attracting females, which explains the common occurrence of sexual dimorphism (Baylis 1981). In addition, among external fertilizing teleost fishes, males are typically the care-givers, in contrast to other vertebrates, which ensure that they fertilize the eggs and protect them after oviposition. The need for the protection of eggs is reinforced by environmental stochasticity (Baylis 1981). Where the risk of eggs being swept away to unfavourable environments is high, there will be an advantage in holding eggs fast. The combination of external fertilisation, environmental stochasticity, and differential rates of gametogenesis are ultimately what led to the evolution of paternal care in fishes (Baylis 1981). Parental care is far more common in freshwater fish compared to marine fish, occurring in 60% of freshwater families compared to only 16% of marine families (Blumer 1982). This difference is explained by the higher environmental stochasticity of rivers compared to the ocean (Baylis 1981).
The lack of overlap between functional hermaphroditism, parental care, and nest-building caught the attention of researchers and two hypotheses have been advanced to explain this (Kuwamura et al. 2020; Warner and Lejeune 1985). The first hypothesis relates to the size-advantage model (SAM), a frequently quoted model of the evolution of protogyny (female to male sex change) and protandry (male to female sex change). Its central tenet is that sex change occurs when one sex reproduces more efficiently when small and young, and the other sex when large and old (Kuwamura et al. 2020; Warner 1975). Warner and Lejeune (1985) hypothesised that nesting males can spawn with only a limited number of females, which negates an advantage that would accrue to large, old males that spawns pelagically.
The second hypothesis advanced by Kuwamura et al. (2020) suggested that freshwater fishes and benthic egg-layers typically have large eggs, compared to marine broadcast spawners, which may require specialised ovarian development that prevents gonadal transition (Kuwamura et al. 2020).
Two Spondyliosoma species, Spondyliosoma emarginatum (Valenciennes 1830), and Spondyliosoma cantharus (Linnaeus 1758), and the closely related Spicara smaris (Linnaeus 1758), are unusual among fishes in that they combine protogyny with nest-building and paternal care (Fairhurst et al. 2007; Harmelin and Harmelin-Vivien 1976; Van Bruggen 1965). This combination of traits is not known in any of the other 36 seabream (family Sparidae) genera and is exceedingly rare among fishes in general.
Spondyliosoma emarginatum is considered endemic to southern Africa and is widely distributed around the shallow continental shelf. It is a small-bodied (maximum length = 312 mm FL), short-lived (maximum age = 8 years) species with early maturation (age at 50% maturity = 3 years), and sex change (age at 50% sex change = 5 years) (Fairhurst et al. 2007). The females have one of the highest annual fecundities among seabreams, whereas the male gonado-somatic index (GSI) is among the lowest (Attwood and Ensair 2020). Its temperate coastal habitat can be defined as stochastic and geographically diverse. During the breeding season, S. emarginatum is sexually dichromatic (unusual among the Sparidae) and the males express territorial and nesting behaviour (Mann 2013; Penrith 1972).
The nesting behaviour of S. emarginatum in an aquarium was briefly documented by Van Bruggen (1965) and prompted our search for the nests in the wild. No scientific account of the nesting behaviour in the wild has been published for either Spondyliosoma species, although a citizen science project documented the nesting of S. cantharus in the English Channel (Doggett 2015). The nests of Spicara sp. have been described in the Mediterranean (Harmelin and Harmelin-Vivien 1976).
In this paper, we report on an intensive nest survey and video-monitoring of the nesting and spawning behaviour of S. emarginatum. To understand the investments and risks involved in a combination of protogyny, nest-building, and paternal care, a thorough description of the reproduction strategy is necessary. Although paternal care of eggs is suspected and inferred, we have no proof of this practice in the wild for S. emarginatum.
We located a S. emarginatum nesting site and monitored nest-building and paternal care. We describe and quantify the activity of males prior to egg deposition and during egg development, and document the presence and activities of females, other fish species, and invertebrates insofar as they affect the nesting process and investment in reproduction. We were interested in the challenges that nest builders face in a space-limited coastal setting, rich in invertebrate and vertebrate predators. Evidence is presented to speculate on the scale of investment in paternal care, in relation to their short life-expectancy, high prior ovarian investment and the applicability of the SAM.
Materials and methods
Study site and species
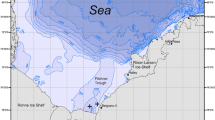
This study was conducted between the 26th of June and the 5th of November 2019 in the north-western region of False Bay, on the southwest coast of South Africa (34° 15′ S, 18° 44′ W) (Fig. 1). Sea surface temperatures in the north-western part of the bay average at 18.8 °C in summer and 15.0 °C in winter (Dufois and Rouault 2012). This region is exposed to lower wave action compared to the rest of the bay (Pfaff et al. 2019; Terhorst 1987) and experiences a semi-diurnal upper microtidal (< 2 m tidal range) environment, which means that tidal driven currents are weak (Terhorst 1987).

Map of study site. Red dot denotes the Spondyliosoma emarginatum compound nesting site
Nesting site description
On the 28th of August, during a SCUBA dive, a large compound nesting site with multiple nests was located between 9 and 15 m, at an average depth of 13 m. The site was arranged in clusters (Fig. 2; Fig. S1). The shape of each cluster was resembled to either a triangle or irregular quadrilateral to aid with area measurements. As such, the sides of each cluster were measured using a measuring tape, and labelled a to c (for triangle clusters) and a to d (for quadrilateral clusters). These measurements were used as per Eqs. (1) and (2) (Hess 2012) to first calculate the semi-perimeter of each cluster (s) from which the cluster area can be calculated (A).

Penguin’s eye view of a Spondyliosoma emarginatum nest cluster. Nests are identified by dotted yellow circles. Top right: a close-up view of one nest. It is surrounded by sand, serpent-skinned brittlestars (Ophioderma wahlbergii) and red-chested sea cucumbers (Pseudocnella insolens). See Fig. S1 for images of the remaining clusters
The number of nests per cluster was counted and divided by the area of the cluster to obtain a nest density (nests m−2) per cluster.
Each cluster was filmed (GoPro Hero4, San Mateo, CA, USA) from above by swimming back and forth along a grid pattern at a constant height off the bottom. One frame per second was extracted from each video, and the frames were stitched together using Panorama Stitcher© to build a composite bird’s eye view of each cluster. A scale was not deemed appropriate for the composite because photographs were slightly distorted or adjusted during the stitching process, meaning that the scale in the foreground and background can differ. To compensate for the lack of an overall scale, each picture was labelled with the size of one nest (Fig. 2; Fig. S1).
Individual nest photographs were taken regularly to measure the size of nests. The presence of eggs on each nest was monitored. Every photograph was taken with a camera (GoPro Hero4, San Mateo, CA, USA) at a fixed height off the seabed. A photograph of a 20 by 20 cm quadrat was taken using the same method to later scale the photographs. All nest photographs were processed using ImageJ© (Schneider et al. 2012). For every photograph, the area of each nest was estimated three times by drawing the outline of the nest using the freehand drawing tool. The mean of the three areas was taken as the final nest area.
A CTD (conductivity, temperature, depth) instrument was used to record the bottom temperature on the nesting site during dive operations from the 27th of August to the 6th of September. The CTD was later replaced with an in situ temperature sensor (SBE 37-SMP-ODO, Sea-Bird Scientific), placed on the nesting site. The temperature sensor recorded continuously and operated from the 7th of September until the 10th of October. Hourly wind direction and speed was also provided by the South African Weather Service (SAWS) from the 1st of August to the 31st of December 2019. Weather data were provided by the Strand coastal Automatic Weather Station (− 34.1174°, 18.8124°). Wind direction and speed were used to calculate the north–south wind vector using the equation:
where ns is the north–south wind vector, ws is the wind speed, and wd is the wind direction.
The north–south wind vectors helped classify the strength and types of weather events affecting the nesting site during the nesting period.
In an attempt to separate the duration of activities associated with courtship and nest preparation from paternal care, we used the proportion of nests with eggs observed on each dive to estimate a date at which half of the nests had eggs. We assume that this date represents the average transition from courtship (including nest preparation) to paternal care. We fitted a logistic model to the proportion of nests that had eggs over the development of the nesting season:
where P(D) is the proportion of nests with eggs on Julian day D, D50 is the Julian day at which half of the nests have eggs, and \({\delta }_{D}\) is a measure of the rate of increase in the proportion of nests with eggs. The model was fitted to the data by finding the values of D50 and \({\delta }_{D}\) which minimised the sum of the squared differences between the model prediction and the observed proportion of nests with eggs.
Video deployments
Individual nests were filmed from the 30th of August until the 7th of October 2019 using cameras (GoPro Inc., San Mateo, CA, USA) with waterproof housings fixed onto custom-made stands. Each stand consisted of two perpendicular metal bars, wired to a 1 kg lead weight. The camera mounts were designed to be < 10 cm high in an attempt to limit the intrusiveness to the nesting males. From the start of the study, nests were tagged using numbered plastic labels attached to small lead weights that were placed in the vicinity of the nest. These would allow nests to be identified during the analysis.
To film individual nests, the camera was placed ~ 30 cm from each nest. The presence or absence of eggs on the nest was noted. On subsequent dives, each camera was either moved to another nest or left to film the same nest. On average, three to four cameras were left to film simultaneously. We chose different combinations of nests for filming on each dive, in an attempt to simultaneously capture nests from different clusters and to sample a mix of those with and without eggs. All the footage was captured between 08:00 and 16:00, at a mean time of day of 11:00. Each video length varied from 18 to 157 min, with a mean time of 92 min. See Table S1 for a summary table of all footage captured.
Video analysis
After an initial sorting of the videos, some footage was discarded due to poor quality and bad camera placement (2 out of 24 nests) or nest abandonment (4 out of 24 nests). The suitable 72 videos that remained covered the behaviour of 18 males (100 h of footage). Males were named using three digit numbers from 001 to 018.
Each video was analysed in full using the behavioural analysis software BORIS (Friard and Gamba 2016). BORIS is an open-source software used for event-logging of video or audio recordings. A predetermined set of behaviours was chosen, based on our perceived relevance to the nesting and spawning processes (Table 1). The table of behaviours (ethogram) was loaded into the software and each behaviour was assigned a letter key. Then any time a behaviour was observed on the video, the appropriate key was pressed and the software recorded the given behaviour with a timestamp. BORIS was used only for event logging and all further statistical analyses were carried out in R (R Core Team 2021). To account for any distress caused by camera placement and nearby divers, video analysis started 5 min from when the male first returned to his nest. In addition, behaviours within 3 min prior to and post any diver sightings in the frame were excluded and that time was removed from analysis time. If a male left the frame and failed to return before the end of the video, the end of the analysis was marked as the last shot of the male in frame. The frequency (counts h−1) of each behaviour was measured per video. If a behaviour was absent, it was assigned a value of 0 counts h−1. For some behaviours (i.e., off nest, female on nest, aggressive bouts), the total duration (min h−1) and mean event time (s) of each behaviour were recorded. Similarly to frequency, if a timed behaviour was absent from a video, it was given a duration of 0 min h−1. However, if a behaviour was absent from a video, it did not contribute a mean event time value. In summary, each behaviour in each video has a maximum of three variables: a total frequency (counts h−1) and, when applicable, a total duration (min h−1) and mean event time (s) (Table 1).
Behavioural effect of egg absence or presence on the nest
Descriptive statistics (mean, standard deviation, and coefficient of variation) were calculated for each behaviour variable by averaging across all males (N = 18). Most males were filmed on more than one day, so variables were first averaged across videos per male to get one value per male, to give each male an equal weighting. Sixteen males were filmed in only one egg group, and two were captured with and without eggs. The same descriptive statistics were then calculated by averaging across only males guarding empty nests (n = 5) (no egg group) and then only males guarding nests with eggs (n = 15) (egg group). Only two males were recorded in both groups, and in these cases, the variables were first averaged across videos per egg group for each male.
A nested analysis of variance (ANOVA) design was implemented to compare the difference in behavioural parameters between the egg groups. Egg presence or absence was set as the main effect, and the male identity was set as the nested effect. To comply with the conditions of a nested design, we removed the two males with observations in both groups from the egg group, since the egg group contained more males than the no egg group. For the nested ANOVA, each video was included, without averaging variables across videos per male first as was done for the descriptive statistics. Nested ANOVA assumes equal variance between groups, which was assessed using a Levene’s test for each behavioural variable.
For behavioural variables where Levene’s test revealed heteroscedasticity (p < 0.05), a Welch’s T-test was carried out instead of a nested ANOVA to compare observations between egg groups. The males omitted from the ANOVA were included here since Welsh’s T-tests do not admit nesting effects. Welch’s T-test disregards variation among the males, by lumping all data and using an average value per male.
Detailed recording of behavioural events
Descriptive accounts of some complex behaviours are given, as they could not be adequately represented quantitatively. These included clearing the nest of invertebrates, courtship and spawning events, a multiple male aggressive bout, and a sneaker male event.
Eggs and larvae
During each dive all nests were inspected for eggs. Upon the appearance of eggs on the site, eggs were sampled from 19 nests using small vials. Eggs were collected from the same nests on consecutive days. The maximum number of consecutive days eggs were captured from the same nest was 5 days, and the average was 3 days. Eggs were collected by gently scraping the surface of the nest using a small spoon and guiding the eggs into a plastic vial. The vials were brought back to the boat, and the seawater was immediately injected with formalin to 5% strength. The samples were kept at room temperature until further analysis. The eggs were photographed with a Nikon DS-Fi 1 Digital Camera fitted to a Nikon SMZ1500, stereo dissecting microscope interfacing with NIS Elements Basic Research 3.2 imaging software. The photographs were used to capture the progression of developmental stages, identify different substrata that eggs may attach to, and identify if eggs of different developmental stages were present on one nest at any one time.
Results
Nesting site description
The nests were scattered on and between outcrops of Cape Peninsula granite. Six nest clusters, labelled A to F (Fig. 2; see Fig. S1 for additional cluster images), ranged in size from 8 to 39 m2. Each cluster was within 5 to 10 m from another cluster. The density of nests in each cluster ranged from 0.6 to 1.3 nests m−2. Four of the clusters (A to D) were on the sandy bottom, while the remaining two clusters (E and F) were on separate granite outcrops. The maximum number of nests observed on the site across all clusters was 83. The average nest size was 0.3 m2 (SD = 0.12 m2, n = 39), and ranged from 0.04 to 1.0 m2. The nests were directly bordered by sand and calcareous debris, mainly comprised of mollusc shells. At a distance beyond 30 cm from the nest borders, the seafloor was covered with a variety of ophiuroids, crinoids, holothuroids, sponges, sea fans, and soft corals. It was immediately clear that the fish kept the invertebrates off the nests.
Water temperature on the nests rose from 11 °C on the 27th of August to 16 °C on the 9th of September but stabilised thereafter between 13 and 17 °C (Fig. 3). A 2 °C dip in temperature occurred on the 20th of September and a near 4 °C drop between the 3rd and 5th of October (Fig. 3). These temperature drops were both preceded by a few days of south-easterly gales (Fig. 3). Throughout the nesting period, southerly winds dominated, punctuated by northerly winds associated with coastal lows and fronts. On one occasion, we documented the destruction of nests following a south-easterly gale. Multiple nests had disappeared, and many were covered in sand. In the following days after this event, most of the nests were rebuilt.

North south wind vectors, for which a southerly wind is positive (top), in situ temperature on the nesting site (middle) and percentage of nests with eggs on the nest (bottom). Red circles identify south-easterly gale events, and red arrows identify temperature drops. The last south-easterly event (C) was accompanied by a sudden drop in percentage of nests with eggs. The red line denotes the logistic equation model used to model the percentage of nests with eggs over time. The black cross denotes the date at which 50% of the nests on the site had eggs. The outlier on the 15/10/2019 was omitted from the model. A south-easterly gale had recently occurred, and it was too difficult to tell by looking at the nests if the eggs had been swept away or if they were simply covered with sand
Nests were inspected for eggs from the 28th of August, yet no eggs were observed on any nests until the 3rd of September (Fig. 3). The number of nests with eggs then steadily increased from the 3rd of September until reaching a maximum of 58 nests with eggs on the 24th of October, out of a total of 83 nests. Thereafter, the number of nests with eggs decreased to reach zero on the 5th of November with no further evidence of egg-laying. During the increase, two dips in the number of nests with eggs occurred, one between the 21st and 26th of September and another between the 7th and 16th of October. The number of consecutive days that a nest held eggs ranged from 2 to 9 days.
Fifty-one days elapsed from the first to the last record of eggs on any nest, which represents the length of the spawning season. Halfway into the spawning season, the percentage of nests with eggs remained below 16%. Thereafter, the percentage of nests with eggs increased rapidly to reach a maximum of 72.5%. The spawning season ended (when no nests held eggs) within 12 days of this peak. On the assumption that nesting began on the date of our discovery of the first nests and that it ended on the first day after peak spawning on which no eggs were observed on any nest, it is estimated that the average male spent 77% of the 67-day nesting period engaged in nest-preparation and courtship, and that the remaining 23% was spent on paternal care (Fig. 3).
Behavioural effect of egg absence or presence on the nest
Once a camera was placed and turned on, it took males a mean time of 4 min 28 s (SD = 2 min 59 s, N = 72) to return to the nest. There was no significant effect of video length on the percentage of time each male spent on the nest (mean = 79.4, p = 0.45, R2 = 0.01, df = 70, N = 72).
Five out of 15 behavioural variables were omitted from the nested analysis of variance (ANOVA) due to heteroscedasticity (p < 0.05) (Table S2).
Males remained active throughout nesting and carried out multiple behaviours. The nested ANOVAs revealed that most of the behaviours that were tested decreased in frequency and duration (Table 2) in the presence of eggs. Mean event time for nest departures and time out of frame also decreased in the presence of eggs (Table 2). Results for the ANOVA tests and Welch’s T-tests can be seen in Tables S3 and S4 respectively.
In the presence of eggs, males spend 86% (52 min h−1) of their time on the nest compared to 51% (31 min h−1) in the absence of eggs (p < 0.001) (Fig. 4; Table 2). This increased presence on the nest was accompanied by a reduced frequency of nest departures (p < 0.001) (Fig. 5; Table 2), and a reduction in the length of these departures (p < 0.001) (16 s when guarding eggs compared to 56 s on empty nests) (Fig. S2; Table 2). The apparent purpose of these departures was either to clear the nest, chase away conspecifics and other species, or to take evasive action during a predator swim-by. The length of departures due to predator avoidance averaged at 51 s (SD = 40, N = 30) but in some instances could extend up to 3 min. Swim-bys of likely fish predators captured on video were the Cape fur seal (Arctocephalus pusillus), the white-breasted cormorant (Phalacrocorax lucidus), and the African penguin (Spheniscus demersus).

Boxplot depicting the percentage of time spent on the nest by Spondyliosoma emarginatum males in the absence and presence of eggs on the nest. Significant value reflects the result from the nested analysis of variance (ANOVA). *p < 0.05, **p < 0.01, ***p < 0.001

Boxplots depicting the frequency of Spondyliosoma emarginatum male behaviours in the absence and presence of eggs on the nest. Significant values reflect results from the nested analysis of variance (ANOVA). *p < 0.05, **p < 0.01, ***p < 0.001
In the presence of eggs, males decreased their territorial and aggressive behaviours. They dropped the frequency and duration (Table 2) of aggressive bouts and the frequency of chases towards conspecifics (Table 2). These conspecific chases decreased in frequency towards other males but not towards other females (Table 2; Fig. S3). The frequency of conspecific males swimming over nesting males also decreased on nests with eggs (Table 2; Fig. S4).
Courtship displays decreased in frequency in the presence of eggs, occurring on average 9 counts h−1 on empty nests compared to 6 counts h−1 when guarding eggs (Table 2). Nevertheless, courtship display was the behaviour which took up the most total time for males guarding eggs (2.42 min h−1) and the second most time for males on empty nests (0.93 min h−1) (Table 2). Females visited empty male nests more frequently than they visited nests with eggs (Table 2), but the total duration of these visits as well as the average event time per visit remained the same irrespective of egg presence (Table 2).
Some behaviours such as clearing the nest and fanning did not change in frequency before or after eggs. Fanning was the most frequent of all behaviours irrespective of egg presence (82 counts h−1) and was also the most frequent behaviour for males guarding eggs (86 counts h−1) (Table 2). The frequency of cleaning the nest however was the most frequent behaviour for males on empty nests (84 counts h−1) but dropped significantly for males guarding eggs (8 counts h−1) (Table 2). On the 30th of August, one male cleaned his nest by picking at algae growing on the surface at a frequency of 196 counts h−1.
Detailed behavioural descriptions
Clearing the nest
Cleaning and clearing are distinguished from each other in the size of the item cleared and the action of the male. Cleaning involves picking at sand or small pieces of vegetation on the nest and eating or spitting them out straight away. In contrast, clearing involves the male actively picking up and swimming away with the item or organisms to clear it off the nest.
The clearing of unwanted items or organisms from the nest was one of the more captivating behaviours observed. As males cleared their nests by depositing unwanted intruders throughout the site, they often dropped the item into the nest of another male. The most common item cleared was intact gastropod shells. It is highly likely that most of these shells were occupied by hermit crabs, as almost all shells inspected during dives were occupied by hermit crabs, but this was not possible to confirm from the footage. Gastropod shells were the only item cleared significantly more in the presence of eggs (p = 0.05, df = 14.9, t = − 2.11).
Courtship and spawning
Courtship displays involved the male swimming in circles at a tilted angle over the nest surface to display accentuated nuptial dress colours to females swimming above (Fig. 6). Once the female was on the nest and inspecting the nest surface, the male repeatedly swam up and down around her and frequently nuzzled her cloaca. If the female left the nest, the male frequently chased after her. There was one exceptional case where a male (009) successfully attracted three females to his nest at the same time. While males often chased away any other female if a female was present on his nest, this male did not (Fig. 6).

Left: Spondyliosoma emarginatum male in courtship display behaviour. He circles the nest at an angle to show off his nuptial dress colours to a female swimming overhead. Right: Three females inspect the nest of a male (not in frame)
Spawning was captured on three separate nests (Table 3). Two events occurred around 13:00 (male 010 and male 009) on the 7th and 14th of September, whereas the other event happened around 10:30 (male 017) on the 2nd of October. For male 010 and 017, each spawning occurred with a single female who released her eggs in batches. She stayed on the nest for the entire video (56 min). For male 009, it was not possible to confirm if all spawn came from the same female, as the female swam in and out of the frame between spawnings and she had no physical features strong enough to confirm identity.
Multiple male aggression
One multiple male exchange was observed during which some males developed a colouration that differed from the usual black and white bands. The top half of their body was much darker than the bottom half, a colouration that was not observed in any other instance (Fig. S5). In addition, pairs of males seemed to engage in a “duel” in which they would swim in circles parallel to each other and regularly curve their body from snout to tail.
Sneaker male
A sneaker male was observed on the 7th of September between 12:00 and 13:00. A male attempted to sneak into a paired spawning event between a nesting male courting a female from his nest (Fig. 7). As the nesting male is courting the female to his nest, another male, herein referred to as sneaker male, can be observed waiting in the background hiding behind rocks and vegetation. As the male and female join for gamete release, the sneaker male darts in between them. The female dashes off, and the nesting male attacks the sneaker male who also eventually leaves. There is no obvious release of gametes from the footage suggesting the sneaker male was unsuccessful on this occasion.

Sequence of events in a sneaker male intrusion. Panel A A sneaker male hides in the background as a nesting male and female get ready for spawning. Panel B The sneaker male jolts out of his hiding place to attempt a sneak fertilization. Following this, the nesting male bites and chases off the sneaker male whilst the female swims away
Eggs and larvae
Eggs were laid in dense layers over the nests and were mostly directly attached to the nest surface but in some cases, the eggs were also attached to algae growing on or bordering the nest surface. The following egg developmental stages were observed (Fig. S6): blastula, gastrula, and an early and late embryo stages in which the yolk sac was greatly reduced. None of the larvae sampled were in pre-flexion phase, which is commonly characterised by full absorption of the yolk sac and flexion of the notochord. Eggs of different developmental stages were found on the same nest, with an egg in the blastula stage and another in an early larval stage.
Discussion
Male investment in reproduction
A variety of behaviours previously undescribed for Spondyliosoma emarginatum were captured on a nesting site. The videos confirm that males excavate and defend nests and tend to their eggs until hatching. Nesting males limit their activities to nesting, courtship, and egg defence and only spend about 13 min per hour off their nest over the 67-day nesting period. This amount of time spent on the nest is similar to that allocated by nesting Chromis crusma (Valenciennes in Cuvier & Valenciennes 1833) (80%, Navarrete-Fernández et al. 2014) and Micropterus salmoides (Lacepede 1802) (97%, Maxwell et al. 2018).
S. emarginatum males build nests in close proximity to one another and form clusters. This can also be referred to as colonial nesting or fish spawning aggregations (Chérubin et al. 2020). Colonial nesting has been observed in the two other nesting Sparidae species: Spondyliosoma cantharus (Collins and Mallinson 2012) and Spicara smaris (Harmelin and Harmelin-Vivien 1976). Colonial nesting is a common occurrence for nesting fish species (Drazen et al. 2003) and may be a result of predation risk or sexual selection. Aggregations can increase chances of brood survival, as it decreases the chances of any single nest being targeted. For example, in cooperatively breeding cichlids, Neolamprologus pulcher (Trewavas and Poll 1952), individuals invested less in antipredator defences (e.g., alarm calling and vigilance) at high densities (Jungwirth et al. 2015). Nesting aggregations also allow for increased mate-choice by females. S. emarginatum females often swam-by and visited nesting males without spawning and this could be part of a mate selection procedure. In colonial nesting grounds for shell-brooding cichlids, Lamprologus callipterus (Boulanger 1906), females preferred to breed with males surrounded by many neighbours (Schütz et al. 2016). Aggregations allow females to “window-shop” and choose the best males to spawn with. Female mate-choice in protogynous species has been shown to result in high levels of variance in reproductive success among males (Benvenuto et al. 2017).
The cost of reproduction to males is a combination of gonadal investment, energy-sapping nesting activities, lost feeding opportunity and the heightened risk of predation. Sperm production is not the major cost. For an average-sized ripe male (300 g) the GSI (1.8%) indicates that approximately 5.6 g mass is lost to sperm, whereas the total condition loss over the nesting period is in the order of 10% or 30 g body mass (Attwood and Ensair 2020). The video analysis suggests that spawning is infrequent, the number of females fertilised per male is limited to one or a few and that there is no sperm competition, which collectively suggests that the male does not release much sperm.
Nesting activities were sufficiently lengthy and energy-demanding to account for the substantial loss of condition measured in males, a loss that occurs in the quarter after the condition loss in females (Attwood and Ensair 2020). The male’s activities recorded on the nest, and the limited time spent off the nest suggest that they do not feed during the 67-day nesting period. Seabreams typically use body fat for gametogenesis but continued feeding supplements gametogenesis during the spawning season (Aristizábal 2007). We did not observe any male S. emarginatum feed whilst on the nest in any of the videos. If, as we suspect, male S. emarginatum do not feed during nesting, they will have limited capacity to replace their sperm during the season. This hypothesis aligns with our interpretation that very little sperm is needed relative to broadcast spawners.
Ovarian development in most seabreams is asynchronous, and they are batch spawners, releasing small quantities of oocytes over 2 to 6 months (Brouwer and Griffiths 2005; Mann 2013). The spawning season of S. emarginatum (estimated at 51 days on this site in 2019) is considerably shorter than that of most seabreams. The trade-off is that the GSI of ripe females is high, suggesting that oocytes are accumulated and released over a short period. Spawning is more concentrated in S. emarginatum than in the broadcast spawning seabreams. S. emarginatum is closer to a capital spawner (McBride et al. 2015) than other seabreams.
Female S. emarginatum can feed unabated during the spawning season, but the season is probably truncated by the energetics of the male. The energy demand coupled with a lack of nutrition in the nesting male limits the spawning season. The bet-hedging strategy employed by other seabreams, which release their eggs pelagically over a long season, is likely traded off in S. emarginatum for the benefits of paternal care. We suspect that a drastic reduction in egg predation is achieved by keeping eggs in a nest, away from filter-feeders during the endogenous feeding phase, which could boost larval survivorship by orders of magnitude (Popiel et al. 1996; Tyler 1995).
Hatchlings leave the nest to commence exogenous feeding once the yolk sac has been absorbed. Hatching takes between 7 and 10 days from fertilisation (Beckley 1989), which agrees with our longest observed consecutive run of eggs on a nest of 9 days. As we found eggs of different stages in a nest, it is possible that longer runs (> 9 days) may have gone unobserved. The best fit of the logistic model suggest 15 days are spent on the care of eggs. The presence of different stages of eggs allows for the interpretation that S. emarginatum could be polygamous, as we suspect after witnessing several females visiting a nest with eggs in one video. Nevertheless, given that the longest run of eggs on a single nest did not exceed the hatching period, the number of females that mate with any given male is likely to be small.
The trade-off introduced by nesting involves not only the length of the spawning season, but also the number of years over which spawning can take place. Nesting exposes nest-bound males to predation. Although we did not witness predation, males were vigilant and temporarily fled their nest when danger presented. We speculate that the unusually short lifespan of S. emarginatum among seabreams is a consequence of heightened exposure to predation. This must be considered as a cost in Darwinian rather than direct energetic terms. The timing of the switch from female to male in protogynous seabreams is mediated by the adult mortality rate, to maintain an optimal sex ratio (Buxton and Garratt 1990; Kerwath et al. 2008). We hypothesise that predation on males has shortened the lifespan and reduced the ages at maturity and sex-change, in comparison to other seabreams.
Activities on the nest
While many behaviours decreased in frequency due to egg presence, they never completely stopped. Energetically demanding behaviours such as fanning and clearing (Jones and Reynolds 1999; van Iersel 1953) were continuous throughout the nesting period. In the presence of eggs, clearing keeps egg predators away whereas fanning keeps the eggs well ventilated and removes metabolic waste and silt or sand build up (Wootton and Smith 2014). The fanning rate increases with brood size and correlates with weight loss in bluegill sunfish Lepomis macrochirus (Rafinesque 1819) (Coleman and Fischer 1991). Experiments on three-spined stickleback Gasterosteus aculeatus (Linnaeus 1758) also revealed that parental activities (with a focus on fanning) caused an expenditure rate of 12.3 J g−1 h−1, which was significantly higher than the expenditure of 3.9 J g−1 h−1 for non-parental males (Smith and Wootton 1999). Fanning occurred at similar rates throughout the entire nesting period for S. emarginatum irrespective of egg presence suggests a large energetic expenditure throughout the nesting period and is likely to be an important limiting factor for brood size and recurrent fertilisations during a season. The uninterrupted rate of fanning throughout the nesting period also implies that fanning serves a dual purpose for S. emarginatum males. Prior to oviposition, fanning helps keep the nest clear of debris and invertebrates. Following oviposition fanning then helps keep the eggs ventilated and clear of debris.
Clearing was regular throughout the nesting period irrespective of egg presence, but the types of items cleared did change. Hermit crabs were cleared significantly more in the presence of eggs, which suggest they may be an important egg predator at this site. No other invertebrate was removed more often in the presence of eggs; however, it is also possible that regular fanning is enough to keep lightweight predators off, such as red-chested sea cucumbers Pseudocnella insolens, cask sea cucumbers Pseudocnella sykion, and serpent-skinned brittlestars Ophioderma wahlbergii. Some large invertebrate predators, notably the 20 cm diameter starfish Marthasterias glacialis (Clark Hubert Lyman 1923), required considerable effort for males to lift from the nest and move away.
On several instances, conspecifics swam over nesting males and even came to inspect the nest surface when the nesting male was gone. Conspecific predation also occurs in other nest-tending species (Gross and MacMillan 1981; Nakazono et al. 1989; Sikkel 1998). For example, in nesting Bluegill Sunfish L. macrochirus, predation is mainly by conspecifics (Gross and MacMillan 1981). Predation pressures have helped explain the benefits of colonial nesting since larger numbers help alleviate the pressure of predation through collective defence or selfish herd effects (Gross and MacMillan 1981). Predation increases the necessity for males to remain on their nest to avoid brood predation and adds to the energetic costs spent during nesting. Predation pressures may also lead to females preferring to lay eggs on nests which already have eggs. All spawning events captured in this study were by a female laying her eggs with a male currently guarding some eggs. Female preference for laying eggs among other eggs has been documented in other species such as Hypsypops rubicundus (Girard 1854) (Sikkel 1989) and is thought to lead to increased egg survival (Jamieson 1995). Alternatively, it could also be that since the observed spawning events occurred towards the end of the spawning season, the majority of the nests already had eggs. An additional find about conspecifics swimming over nesting males is that the rate of swim-bys decreased towards the end of the nesting period. This coincided with fewer males chasing away conspecifics. It is likely that since the number of males busy guarding eggs increased as the nesting period progressed, fewer males remained swimming over the nesting site.
These energetic demands help explain sneaker males, who, when successful, can offload the energetic costs of nest-tending and guarding of their offspring to other males. When this event occurs in nests, it is known as cuckoldery (Alonzo and Heckman 2010; DeWoody and Avise 2001). Until now, sneaker males in the Sparidae are mentioned only for the broadcast-spawning Australasian snapper Chrysophrys auratus (Forster 1801) (Smith 1986) and Santer seabream Cheimerius nufar (Valenciennes 1830) (Garratt 1991). Both accounts are in aquaria and describe similar events: as a female swam along the surface of the tank, she was trailed by several males and when she released her eggs, several males released their sperm. These events are more appropriately described as sperm competition in a spawning aggregation, where males compete for a female. Sneaker males require a dominant male and other satellite males which does not seem to be the case in these accounts. The S. emarginatum sneaker event captured is likely the first account of sneaker males and cuckoldery in the wild for a seabream.
Life-history strategy
Despite the rarity of the combination, this nesting strategy is compatible with a protogynous life-history, since larger individuals will be more capable of excavating and guarding a nest and should be less prone to predation. Warner and Lejeune’s (1985) statement that nest-building, and paternal care limits the number of females that a large male can fertilise is relevant here, but we suggest that the size advantage lies not in the number of females that get fertilised, but rather the probability of maintaining a nest and defending eggs until hatching in a competitor- and predator-rich setting. Importantly they will have greater stamina to prolong the nesting period. Nesting behaviour and paternal care may provide the size-advantage for males needed to meet Warner’s requirement for protogyny (Warner 1975).
S. emarginatum is dichromatic, a rare trait among Sparidae species, which is likely associated with the nesting habitat (Attwood and Ensair 2020). Dichromatism is a sign of sexual selection (Darwin 1875; Mank et al. 2005) and points to the type of reproductive competition among males (Mank et al. 2005). This competition may come from a scarcity of suitable habitats (nests in this case), a scarcity of females, or both. The male to female sex ratio for S. emarginatum and S. cantharus is 4.2:1 and 4.1:1 respectively (Fairhurst et al. 2007; Gonçalves and Erzini 2008). The other Sparidae nesting species, S. smaris, has an identical female to male sex ratio of 4.1:1 (Dulčić et al. 2003). We suspect that the sex-ratio is self-regulating in these protogynous seabreams (Buxton and Garratt 1990; Kerwath et al. 2008; Tunley et al. 2009) and that this asymmetrical ratio is optimal for this reproductive strategy, which requires less sperm than is the case in other seabreams.
The Spondyliosoma life-history strategy can be described as opportunistic, as the generation time is short (Winemiller 1992; Winemiller and Rose 1992). Spondyliosoma has evolved life-history traits that allow for “efficient recolonization of habitats over relatively small spatial scales” (Winemiller and Rose 1992). By contrast, most Sparidae can be described as periodic strategists, with long generation times. Selection pressures resulting in an opportunistic strategy come from stochastic environments that change frequently over small spatio-temporal scales. S. emarginatum is one of the most abundant species among South Africa’s 42 seabreams, and its success might be attributed to its ability to flourish in stochastic conditions and in a variety of ecosystem types. The short-term stochasticity of the environment was highlighted in this study when strong south-easterly gales led to the destruction of multiple nests. Despite being a benthic egg layer, the Spondyliosoma spp. complex is more widespread than other seabream genera, as it ranges from the North Atlantic off the western British Isles to Durban in the south-western Indian Ocean. In so doing it spans a massive range of habitat and biogeographic zones. This is very rare for a protogynous species, as a recent review stated that only 5% of protogynous species have adapted to temperate environments (Pla et al. 2021).
The opportunistic strategy is possibly what has allowed Spondyliosoma spp. to occupy widely divergent coastal environments, in both hemispheres, spanning 86 degrees of latitude. Whereas fish species resilience has frequently been linked to life-history combinations (King and McFarlane 2003), we contend that the natural ranges of fishes might be similarly affected. Indeed, life-history is a predictor of invasive potential (Capellini et al. 2015; Marchetti et al. 2004).
Conclusions
The size-advantage model (SAM) applies differently to S. emarginatum than it does to pelagic spawning seabreams. For this nest-building protogynous fish, the advantage of a large male is its increased stamina for nest-building, nest maintenance, and nest-guarding, yet they spawn with a limited number of females. In contrast, large protogynous pelagic spawners have the advantage of a greater access to females.
Spondyliosoma spp. differ from other seabreams in that they are short-lived opportunists. Predation may lead to short longevity, early maturation, and as a consequence small size. Their strategy hinges on reducing mortality in the younger stages (from eggs to hatching), thereby removing the necessity for extreme bet-hedging (high longevity, high life-time fecundity, long spawning seasons) in variable coastal environments.
The fact that these benthic egg-laying seabreams are more widely distributed than their pelagic broadcast-spawning counterparts is a paradox that might be explained by their opportunistic life-history and begs an investigation into a correlation of life-history characteristics and range restriction within the family.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Alonzo SH, Heckman KL (2010) The unexpected but understandable dynamics of mating, paternity and paternal care in the ocellated wrasse. Proc Roy Soc B: Biol Sci 277:115–122. https://doi.org/10.1098/rspb.2009.1425
Aristizabal EO (2007) Energy investment in the annual reproduction cycle of female red porgy, Pagrus pagrus (L.). Mar Biol 152:713–724. https://doi.org/10.1007/s00227-007-0729-6
Attwood CG, Ensair HAM (2020) Life-history trade-offs among four sympatric seabreams. Afr J Mar Sci 43:323–337. https://doi.org/10.2989/1814232X.2020.1794957
Bateman AJ (1948) Intra-sexual selection in Drosophila. Heredity 2(3):349–368. https://doi.org/10.1038/hdy.1948.21
Baylis JR (1981) The evolution of parental care in fishes, with reference to Darwin’s rule of male sexual selection. Environ Biol Fish 6:223–251
Beckley LE (1989) Larval development of Spondyliosoma emarginatimi (Cuvier & Valenciennes) (Pisces: Sparidae) from southern Africa South. Afr J Zool 24:187–192. https://doi.org/10.1080/02541858.1989.11448151
Benvenuto C, Coscia I, Chopelet J, Sala-Bozano M, Mariani S (2017) Ecological and evolutionary consequences of alternative sex-change pathways in fish. Sci Rep 7:9084. https://www.nature.com/articles/s41598-017-09298-8
Blumer LS (1982) A bibliography and categorization of bony fishes exhibiting parental care. Zool J Linn Soc 75:1–22. https://doi.org/10.1111/j.1096-3642.1982.tb01939.x
Brouwer SL, Griffiths MH (2005) Reproductive biology of carpenter seabream (Argyrozona argyrozona) (Pisces: Sparidae) in a marine protected area. Fish Bull 103:258–269
Buxton CD, Garratt PA (1990) Alternative reproductive styles in seabreams (Pisces : Sparidae). Environ Biol Fishes 28:113–124. https://doi.org/10.1007/BF00751031
Capellini I, Baker J, Allen WL, Street SE, Venditti C (2015) The role of life history traits in mammalian invasion success. Ecol Lett 18:1099–1107. https://doi.org/10.1111/ele.12493
Chérubin LM, Dalgleish F, Ibrahim AK, Schärer-Umpierre M, Nemeth RS, Matthews A, Appeldoorn R (2020) Fish spawning aggregations dynamics as inferred from a novel, persistent presence robotic approach. Front Mar Sci 6:779. https://doi.org/10.3389/fmars.2019.00779
Clark Hubert Lyman (1923) The echinoderm fauna of South Africa. Annals of the South African Museum. Annale van die Suid-Afrikaanse Museum., 13:221–435. Cape Town :South African Museum,1898–2004. https://www.biodiversitylibrary.org/part/33491
Coleman RM, Fischer RU (1991) Brood size, male fanning effort and the energetics of a nonshareable parental investment in bluegill sunfish, Lepomis macrochirus (Teleostei: Centrarchidae). Ethology 87:177–188. https://doi.org/10.1111/j.1439-0310.1991.tb00245.x
Collins KJ, Mallinson JJ (2012) Surveying black bream, Spondyliosoma cantharus (L.), nesting sites using sidescan sonar. Underw Technol 30:183–188. https://doi.org/10.3723/ut.30.183
Darwin C (1875) The descent of man, and selection in relation to sex. Nature 11:305
De Mitcheson YS, Liu M (2008) Functional hermaphroditism in teleosts. Fish Fish 9:1–43. https://doi.org/10.1111/j.1467-2979.2007.00266.x
DeWoody JA, Avise JC (2001) Genetic perspectives on the natural history of fish mating systems. J Hered 92:167–172. https://doi.org/10.1093/jhered/92.2.167
Doggett M (2015) The Black Bream project. http://www.mattdoggett.com/the-black-bream-project/
Drazen JC, Goffredi SK, Schlining B, Stakes DS (2003) Aggregations of egg-brooding deep-sea fish and cephalopods on the gorda escarpment: a reproductive hot spot. Biol Bull 205:1–7. https://doi.org/10.2307/1543439
Dufois F, Rouault M (2012) Sea surface temperature in False Bay (South Africa): towards a better understanding of its seasonal and inter-annual variability. Cont Shelf Res 43:24–35. https://doi.org/10.1016/j.csr.2012.04.009
Dulčić J, Pallaoro A, Cetinić P, Kraljević M, Soldo A, Jardas I (2003) Age, growth and mortality of picarel, Spicara smaris L. (Pisces: Centracanthidae), from the eastern Adriatic (Croatian coast). J Appl Ichthyol 19:10–14. https://doi.org/10.1046/j.1439-0426.2003.00345.x
Fairhurst L, Attwood CG, Durholtz MD, Moloney CL (2007) Life history of the steentjie Spondyliosoma emarginatum (Cuvier 1830) in Langebaan Lagoon, South Africa. Afr J Mar Sci 29:79–92. https://doi.org/10.2989/AJMS.2007.29.1.7.71
Friard O, Gamba M (2016) BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol Evol 7:1325–1330. https://doi.org/10.1111/2041-210X.12584
Garratt PA (1991) Spawning behaviour of Cheimerius nufar in captivity. Environ Biol Fishes 31:345–353. https://doi.org/10.1007/BF00002358
Gonçalves JMS, Erzini K (2008) The reproductive biology of Spondyliosoma cantharus (L.) from the SW Coast of Portugal. Sci Mar 64:403–411. https://doi.org/10.3989/scimar.2000.64n4403
Gross MR, MacMillan AM (1981) Predation and the evolution of colonial nesting in bluegill sunfish (Lepomis macrochirus). Behav Ecol Sociobiol 8:163–174. https://doi.org/10.1007/BF00299826
Harmelin JG, Harmelin-Vivien ML (1976) Observations “in situ” des aires de ponte de Spicara smaris (L.) (Pisces, Perciformes, Centracanthidae) dans les eaux de Port-Cros. Trav Sci Parc Natl Port-Cros 2:115–120
Hess A (2012) A highway from Heron to Brahmagupta. Forum Geometricorum 12:191–192
Jamieson I (1995) Do female fish prefer to spawn in nests with eggs for reasons of mate choice copying or egg survival? Am Nat 145:824–832. https://doi.org/10.1086/285770
Jones JC, Reynolds JD (1999) Costs of egg ventilation for male common gobies breeding in conditions of low dissolved oxygen. Anim Behav 57:181–188. https://doi.org/10.1006/anbe.1998.0939
Jungwirth A, Josi D, Walker J, Taborsky M (2015) Benefits of coloniality: communal defence saves anti-predator effort in cooperative breeders. Funct Ecol 29:1218–1224. https://doi.org/10.1111/1365-2435.12430
Kerwath SE, Götz A, Attwood CG, Sauer WHH (2008) The effect of marine protected areas on an exploited population of sex-changing temperate reef fish: an individual-based model. Afr J Mar Sci 30:337–350. https://doi.org/10.2989/AJMS.2008.30.2.10.560
King JR, McFarlane GA (2003) Marine fish life history strategies: applications to fishery management. Fish Manage Ecol 10:249–264. https://doi.org/10.1046/j.1365-2400.2003.00359.x
Kuwamura T, Sunobe T, Sakai Y, Kadota T, Sawada K (2020) Hermaphroditism in fishes: an annotated list of species, phylogeny, and mating system. Ichthyol Res 67:341–360. https://doi.org/10.1007/s10228-020-00754-6
Mank JE, Promislow DEL, Avise JC (2005) Phylogenetic perspectives in the evolution of parental care in ray-finned fishes. Evolution 59:1570. https://doi.org/10.1111/j.0014-3820.2005.tb01806.x
Mann B (2013) Southern African marine linefish species profiles. Special publication, 9.
Marchetti MP, Moyle PB, Levine R (2004) Invasive species profiling? Exploring the characteristics of non-native fishes across invasion stages in California. Freshw Biol 49(5):646–661. https://doi.org/10.1111/j.1365-2427.2004.01202.x
Maxwell RJ, Zolderdo AJ, de Bruijn R, Brownscombe JW, Staaterman E, Gallagher AJ, Cooke SJ (2018) Does motor noise from recreational boats alter parental care behaviour of a nesting freshwater fish? Aquat Conserv Mar Freshwat Ecosyst 28:969–978. https://doi.org/10.1002/aqc.2915
Mcbride RS, Somarakis S, Fitzhugh GR, Albert A, Yaragina NA, Wuenschel MJ, Alonso-Fernández A et al (2015) Energy acquisition and allocation to egg production in relation to fish reproductive strategies. Fish Fish 16:23–57. https://doi.org/10.1111/faf.12043
Nakazono A, Hamada H, Sakurai M (1989) Predation on eggs by conspecific males in two Japanese damselfishes, Pomacentrus nagasakiensis and Chromis notatus notatus, after removal of egg-guarding males. J Ethol 7:97–104. https://doi.org/10.1007/BF02350030
Navarrete-Fernández T, Landaeta MF, Bustos CA, Pérez-Matus A (2014) Nest building and description of parental care behavior in a temperate reef fish, Chromis crusma (Pisces: Pomacentridae). Rev Chil Hist Nat 87:1–9. https://doi.org/10.1186/s40693-014-0030-2
Penrith MJ (1972) The behaviour of reef-dwelling sparid fishes. Zoologica Africana 7:43–48. https://doi.org/10.1080/00445096.1972.11447428
Pfaff MC, Logston RC, Raemaekers SJPN, Hermes JC, Blamey LK, Cawthra HC, Colenbrander DR et a. (2019) A synthesis of three decades of socio-ecological change in False Bay, South Africa: setting the scene for multidisciplinary research and management. Elementa, 7. https://doi.org/10.1525/elementa.367
Pla S, Benvenuto C, Capellini I, Piferrer F (2020) A phylogenetic comparative analysis on the evolution of sequential hermaphroditism in seabreams (Teleostei: Sparidae). Sci Rep 10:1–12. https://doi.org/10.1038/s41598-020-60376-w
Pla S, Maynou F, Piferrer F (2021) Hermaphroditism in fish: incidence, distribution and associations with abiotic environmental factors. Rev Fish Biol Fisheries 31:935–955. https://doi.org/10.1007/s11160-021-09681-9
Popiel SA, Pérez-Fuentetaja A, Mcqueen DJ, Collins NC (1996) Determinants of nesting success in the pumpkinseed (Lepomis gibbosus): a comparison of two populations under different risks from predation. Copeia 1996:649–656. https://doi.org/10.2307/1447529
R Core Team (2021). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH Image to ImageJ: 25 years of image analysis. Nat Methods 9(7):671–675. https://doi.org/10.1038/nmeth.2089
Schütz D, Wirtz Ocana S, Maan ME, Taborsky M (2016) Sexual selection promotes colonial breeding in shell-brooding cichlid fish. Anim Behav 112:153–161. https://doi.org/10.1016/j.anbehav.2015.11.022
Sikkel PC (1989) Egg presence and developmental stage influence spawning-site choice by female garibaldi. Anim Behav 38:447–456. https://doi.org/10.1016/S0003-3472(89)80038-7
Sikkel PC (1998) Competitor intrusions and mate-search tactics in a territorial marine fish. Behav Ecol 9:439–444. https://doi.org/10.1093/beheco/9.5.439
Smith PJ (1986) Spawning behaviour of snapper, Chrysophrys auratus, in captivity (note). NZ J Mar Freshwat Res 20:513–515. https://doi.org/10.1080/00288330.1986.9516170
Smith C, Wootton RJ (1999) Parental energy expenditure of the male three-spined stickleback. J Fish Biol 54:1132–1136. https://doi.org/10.1111/j.1095-8649.1999.tb00866.x
Terhorst A (1987) The seafloor environment off Simon’s Town in False Bay, revealed by side-scan sonar, bottom sampling, diver observation and underwater photography. Dissertation, University of Cape Town
Trivers R (1972) Parental investment and sexual selection
Tunley KL, Attwood CG, Moloney CL, Fairhurst L (2009) Variation in population structure and life-history parameters of steentjies Spondyliosoma emarginatum: effects of exploitation and biogeography. Afr J Mar Sci 31:133–143. https://doi.org/10.2989/AJMS.2009.31.2.2.874
Tyler WA (1995) The adaptive significance of colonial nesting in a coral-reef fish. Anim Behav 49:949–966. https://doi.org/10.1006/anbe.1995.0125
Van Bruggen AC (1965) Records and observations in the Port Elizabeth Oceanarium in 1960. Zool Gart Lpz 31:184–202
van Iersel JJA (1953) An analysis of the parental behaviour of the male three-spined stickleback (Gasterosteus aculeatus L.). Behaviour 3:1–159. http://www.jstor.org/stable/30039128
Warner RR, Lejeune P (1985) Sex change limited by paternal care: a test using four Mediterranean labrid fishes, genus Symphodus. Mar Biol 87:89–99. https://doi.org/10.1007/BF00397010
Warner, RR (1975) The adaptive significance of sequential hermaphroditism in animals. Am Nat 109:61–82. https://doi.org/10.1086/282974
Winemiller KO (1992) Life-history strategies and the effectiveness of sexual selection. Oikos 63:318. https://doi.org/10.2307/3545395
Winemiller KO, Rose KA (1992) Patterns of life-history diversification in North American fishes: implications for population regulation. Can J Fish Aquat Sci 49:2196–2218. https://doi.org/10.1139/f92-242
Wootton RJ, Smith C (2014) Reproductive biology of teleost fishes. Wiley Blackwell
Acknowledgements
We thank the many divers that helped make this study possible, particularly Andrea Plos for providing surface support and dive supervision throughout this project. We thank Sea Technology Services for lending us the temperature and oxygen sensors. We also thank David Smith for helping with some of the dive trips.
Funding
Funding was provided by the South African National Research Foundation (NRF).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethics approval
All ethics were approved by the Science Faculty Animal Ethics committee (SFAEC) at the University of Cape Town, and the study was performed in accordance with the ethical standards as laid down in the 1964 Declaration of Helsinki and its later amendments.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Faure-Beaulieu, N., Attwood, C.G. Remote video analysis of an unusual fish life-history combination: nest-building, paternal care, and protogyny in a seabream. Environ Biol Fish 105, 933–951 (2022). https://doi.org/10.1007/s10641-022-01303-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-022-01303-y