
Abstract
In environmental impact studies conducted in coastal regions, the survey of biodiversity is crucial in areas that are subject to interventions. As the characterization of aquatic invertebrates may indicate important issues that need to be addressed. In this regard, hydrozoans (Class Hydrozoa, Phylum Cnidaria) are recognised as important bioindicators. These organisms are characterized by biradial, tetramerous, or polymerous symmetry, a simple gastrovascular system, and non-cellular mesoglea. The aim of the present study was to perform the first descriptive survey of hydrozoan species found in the estuary of the International Minho River (shared by Portugal and Spain), which is classified as an Important Bird Area and is a site of the Natura 2000 network. Specimens were collected from the tidal marine, brackish and tidal freshwater wetland zones through 1) glass eel fishing bycatch using stow nets at flood tides around the new moon (length of float lines: 10 m; bottom anchored lead line: 15 m; height: 8 m; mesh size: 1–2 mm); 2) beam trawl fishing (bottom trawling for 10 min), including species associated with bivalves; 3) Van Veen grab sampler for sediment sampling; 4) on wood fragments in the saltmarsh of the Coura River (tributary of the Minho River); and 5) individuals on the leaves of aquatic plants. A total of 19 species were identified, four of which were first occurrences for Portugal, six for the mainland coast of the country, and one for Southern Europe [Diphasia fallax (Johnston, 1847)]. Three neotypes are also proposed for the species Hydra viridissima, Hydra oligactis and Gymnangium montagui based on the criteria of the International Commission on Zoological Nomenclature. As these organisms are considered bioindicators, the diversity of the hydrozoan community in the Minho River may indicate good water quality.
Similar content being viewed by others

Introduction
Aquatic ecological assessments are essential to determining future impacts caused by human actions. Thus, biodiversity surveys conducted by taxonomists and environmental specialists are important and enable reliable results on species inhabiting a given location. Hydrozoans (Class Hydrozoa, Phylum Cnidaria) constitute one of the most representative aquatic groups and are recognised as important bioindicators. These organisms are characterized by biradial, tetramerous, or polymerous symmetry, a simple gastrovascular system, and non-cellular mesoglea and are mainly dioecious, generally producing gametes of ectodermal origin (Bouillon et al. 2006). Hydrozoans are broadly distributed in all biogeographical regions (Cornelius 1992a), mainly occupying marine environments, but can also be found in brackish and freshwater ecosystems (Encyclopædia Britannica 2023). The hydrozoan lifecycle involves three stages: free-living planula larvae, sessile colonial polyps, and dioecious sexually-reproducing medusae (Hammond 2009).
Currently, ca. 3,700 to 3,800 valid species are known (Schuchert 2022; Encyclopædia Britannica 2023) nested in two monophyletic groups: Hydroidolina Collins, 2000 and Trachylinae Haeckel, 1879 (Daly et al. 2007). Hydroidolinans comprise three orders: Anthoathecata Cornelius, 1992a, b (hydrozoans without hydrotheca or gonotheca, including aplanulates, capitates, and filiferans), Leptothecata Cornelius, 1992a, b (with hydrotheca and gonotheca), and Siphonophorae Eschscholtz, 1829 (holopelagic with a polymorphic colonial arrangement) (Daly et al. 2007; Hayward and Ryland 2017). This paper presents the first taxonomical survey of hydrozoans sampled from the estuarine zone of the International Minho River, Iberian Peninsula, providing descriptions, type and nomenclatural information, overall distribution and distribution in Portugal, and ecological notes. A total of 19 species were identified, 18 of which are new records for the main section of the river and one for the saltmarsh area of the Coura River (Cordylophora caspia (Pallas, 1771)), four for Portugal (Hydra viridissima Pallas, 1766; Hydra oligactis Pallas 1766, Diphasia fallax (Johnston, 1847), and Sertularia cupressina Linnaeus, 1758), six for mainland Portugal (H. viridissima, H. oligactis, Gonothyraea loveni (Allman, 1859), Obelia dichotoma (Linnaeus, 1758), D. fallax, and S. cupressina), and one for Southern Europe (D. fallax). The establishment of three neotypes is also proposed based on Article 75 and the qualifying conditions of the International Commission on Zoological Nomenclature (ICZN), 1999).
Material and methods
Sampling and preservation
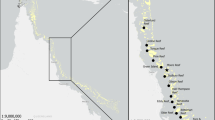
All organisms analysed were sampled in previous campaigns in the marine and brackish tidal zones and tidal freshwater wetland of the Minho River on the Iberian Peninsula in Southern Europe (Fig. 1) using the following methods: 1) on the surface of Egeria densa Planchon, 1846 leaves in Vila Nova de Cerveira (41º56′20.15″N / 8º44′59.43″W); 2) glass eel fishing bycatch in Caminha (41º52′59.00″N / 8º50′14.00″W) using stow nets at flood tides around new moon (length of float lines: 10 m; bottom anchored lead line: 15 m; height: 8 m; mesh size: 1–2 mm) in May 2020; 3) glass eel fishing bycatch in Caminha (41º52′44.80″N / 8º50′26.25″W) in March 2021; 4) glass eel fishing bycatch in La Guardia (41°52′17.43″N / 8°52′8.01″W) in December 2021; 5) beam trawl in Caminha (41°52′04.80″N / 8°51′18.80″W) in September 2021; 6) on sediment samples collected with a Van Veen grab sampler in Valença (42º00′58.20″N / 8º39′29.09″W) in September 2020; and 7) on wood fragments sampled by hand in the saltmarsh area of the Coura River (41°52′41.85″N / 8°50′4.77″W) in January 2022.

Study area in section of International Minho River Basin, Iberian Peninsula, Atlantic Europe
Sampled hydrozoans were fixed and stored in 70% alcohol and deposited in the Natural History Museum of the Iberian Peninsula (NatMIP), Vila Nova de Cerveira, Portugal.
Data from scientific collections
We use the following acronyms and abbreviations for museums and scientific collections: 1) Calder’s manuscript (Calder 2019a), via GBIF (Calder 2019b); 2) COAST (based on the article by Humara-Gil and Cruz-Gómez (2018)); 3) LINN (collections of The Linnean Society of London); 4) MCZ-IZ (Invertebrate Zoology, Museum of Comparative Zoology, Harvard University); 5) NHMD (Natural History Museum of Denmark, also known as Zoological Museum of the University of Copenhagen (ZMUC) or Statens Naturhistoriske Museum (SNM)); 6) NHMUK (Natural History Museum, London, United Kingdom, also known as British Museum (Natural History)—BM(NH)); 7) NMV-OZCAM (National Museum of Victoria, Online Zoological Collections of Australian Museums); 8) RBCM (Royal British Columbia Museum); 9) USNM (Smithsonian Institution, National Museum of Natural History (former abbreviation: NMNH)); 10) ZMA (Zoological Museum Amsterdam, also known as Naturalis Biodiversity Center, Leiden, Netherlands; National Museum of Natural History, Naturalis in Leiden; and RMNH—Rijksmuseum voor Natuurlijke Historie); 11) ZMB (Museum für Naturkunde Berlin-Zoological Collections, also known as Zoologisches Museum der Humboldt-Universität zu Berlin).
Taxonomy, distribution, and ecological notes
Hydrozoans were identified and photographed with a Leica EZ4W stereomicroscope, Nikon Eclipse 50i microscope, Nikon SMZ800 stereomicroscope, and Nikon Digital Sight D5-L1 camera. The following manuscripts were used for information on taxonomy and ecological notes: Synopsis of the British Fauna (Cornelius 1995; Schuchert 2012), Handbook of the marine fauna of North-West Europe (Hayward and Ryland 2017), Portuguese manuscripts (Nobre 1903a, b, 1937; Da Cunha 1940, 1944; Moura 2011; Moura et al. 2012a, b), and additional manuscripts (Cornelius 1979; Schuchert 2004, 2010). The organization of the taxonomic groups and information on synonyms were sought through the website of the World Register of Marine Species (WoRMS Editorial Board 2023). Type information, when available, was primarily based on the Linnean collections (The Linnean Society 2023), Global Biodiversity Information Facility (GBIF 2023), original manuscripts, and related papers.
Results
Taxonomic approach
A total of 19 species were identified, 18 of which constitute new records for the main section of the river and one for the saltmarsh area of the Coura River (Cordylophora caspia (Pallas, 1771)). Four of these species are new records for Portugal (Hydra viridissima Pallas, 1766; Hydra oligactis Pallas, 1766, Diphasia fallax (Johnston, 1847), and Sertularia cupressina Linnaeus, 1758), six are new records for mainland Portugal (H. viridissima, H. oligactis, Gonothyraea loveni (Allman, 1859), Obelia dichotoma (Linnaeus, 1758), D. fallax, and S. cupressina), and one is a new record for Southern Europe (D. fallax). The establishment of three neotypes is also proposed based on Article 75 and the qualifying conditions of the International Commission on Zoological Nomenclature (ICZN 1999).
Phylum Cnidaria Hatschek, 1888
Subclass Hydroidolina Collins, 2000
Order Anthoathecata Cornelius, 1992a, b
Family Cordylophoridae von Lendenfeld, 1885
Genus Cordylophora Allman, 1844
Cordylophora caspia (Pallas, 1771) (Fig. 2a).

Hydrozoans collected throughout International Minho River Basin: a. Cordylophora caspia (Pallas 1771); b. Hydra viridissima Pallas 1766; c. Hydra oligactis Pallas 1766 on Egeria densa Planchon, 1846 leaf; d. Gonothyraea loveni (Allman 1859); e. Obelia dichotoma (Linnaeus 1758) on bivalve Cerastoderma edule (Linnaeus 1758); f. Obelia geniculata (Linnaeus 1758). Scales: 500 µm: a, b, d, f; 2 mm: c, e
Synonyms: Tubularia caspia Pallas, 1771 (original designation);
Bimeria baltica Stechow, 1927; Cordylophora albicola Kirchenpauer in Busk, 1861; Cordylophora americana Leidy, 1870; Cordylophora japonica Itô, 1951; Cordylophora lacustris Allman, 1844; Cordylophora lacustris var. otagoensis Fyfe, 1929; Cordylophora otagoensis Fyfe, 1929; Cordylophora whiteleggi von Lendenfeld, 1887; Tubularia cornea Aghardh, 1816.
Type material: Syntypes: ZMB 1473, 5943 (GBIF 2023).
Type locality: Caspian Sea (Pallas 1771; Schuchert 2004, 2022; Calder 2012).
Material examined: Colonies from Coura River Saltmarsh, Minho River Basin, Portugal (41°52′41.85″N / 8°50′4.77″W) on January 18, 2022 (NatMIP-CHAh-0034).
Diagnosis: Monosiphonic randomly branched colonies with sprouts directed upwards, up to 100 mm from meshed stolons; whitish to brownish hydranths with 14 to 27 capillary tentacles. Elliptical-like gonophores enclosed with yellowish perisarc (Schuchert 2004, 2012; Hayward and Ryland 2017). Notes on medusae and reproductive features: Release into planula when temperature reaches 10ºC (Schuchert 2004); medusae Gaussian curve-shaped, with tentacles or oral arms on manubrium (Schuchert 2012); reproductive season during spring in Italy (Gravili et al. 2015).
Geographical distribution: Temperate to subtropical waters (Schuchert 2012, 2022; Calder 2019a).
Distribution in Portugal: Santo André Lagoon, Santiago do Cacém (Correia et al. 2012); Tagus River estuary (Conde et al. 2013); Minho River: upper estuary (Servia et al. 2006) and lower estuary, Coura River Saltmarsh, Minho River Basin (this study).
Ecological notes: Freshwater to brackish environments (Calder 2012), including estuaries and coastal lagoons, intertidal to depth of 85 m (Schuchert 2004, 2012); can be found near aquatic plants (Pallas 1771), on rocks (Teissier 1965), under stones, on molluscs, such as barnacles (Gravili et al. 2015) and bivalves Dreissena spp. (Schuchert 2012), on polychaete tubes of Ficopomatus enigmaticus (Fauvel, 1923), plastic (Gravili et al. 2015), and wood fragments (this study). Is considered an exotic species (Conde et al. 2013).
Family Hydridae Dana, 1846
Genus Hydra Linnaeus, 1758
Hydra viridissima Pallas, 1766 (Fig. 2b).
Synonyms: Chlorohydra viridissima (Pallas, 1766); Hydra viridis Linnaeus, 1767
Type material: Unknown (GBIF 2023). Thus, we propose here the designation of a neotype in the same geographical type locality (European rivers) because the name-bearing type specimen (i.e. holotype, lectotype, syntype, or prior neotype) is possibly lost/destroyed (Article 75 and qualifying conditions (ICZN 1999)): NatMIP-CHAh-0038 from the International Minho River deposited at the Natural History Museum of the Iberian Peninsula, Vila Nova de Cerveira, Portugal (this study).
Type locality: European rivers (Linnaeus 1767), here designated as International Minho River, Vila Nova de Cerveira, Portugal (this study).
Material examined: Individuals from International Minho River, Vila Nova de Cerveira, Portugal (41º56′20.15″N / 8º44′59.43″W) on May 23, 2022: one designated as neotype (NatMIP-CHAh-0038), and four others (NatMIP-CHAh-0040).
Diagnosis: Solitary freshwater anthoathecatan greenish polyps supported posteriorly by flat plate; hypostome tapered, anteriorly with 4–11 tentacles. Ellipsoid stenotele nematocysts (Schuchert 2012). Notes on medusae and reproductive features: No medusae stage. Hermaphroditism with reproduction vegetative-like, mature eggs globular-shaped; reproductive season in summer (Schuchert 2010, 2012) and spring (this study).
Geographical distribution: Europe (Schuchert 2010, 2012). Other distributions need revision.
Distribution in Portugal: International Minho River, Vila Nova de Cerveira (new record, this study). This is the first taxonomic record of the species for Portugal.
Ecological notes: Freshwater to brackish habitats, mainly in motionless (Schuchert 2010) and less polluted waters (Pallas 1766); symbiotic with green algae Chlorella sp. (Hamada et al. 2020); found on aquatic plants (Schuchert 2012), such as E. densa (this study).
Hydra oligactis Pallas, 1766 (Fig. 2c).
Synonyms: Hydra corala Elrod & Ricker, 1902; Hydra dioecia Downing, 1905; Hydra fusca Linnaeus, 1767; Hydra monoecia Downing, 1900; Hydra pallida Beardsley, 1904; Hydra rhaetica Asper, 1879; Hydra rhistica Asper, 1880; Hydra roeselii Haacke, 1879; Pelmatohydra oligactis (Pallas, 1766).
Type material: Unknown (GBIF 2023). Therefore, we propose here the designation of a neotype in the same geographical type locality (European freshwater environments), based on Article 75 and qualifying conditions (ICZN 1999)): NatMIP-CHAh-0039 from International Minho River, deposited in the Natural History Museum of the Iberian Peninsula, Vila Nova de Cerveira, Portugal (this study).
Type locality: European freshwater environments (Pallas 1766), here designated International Minho River, Vila Nova de Cerveira, Portugal (this study).
Material examined: Individuals from the International Minho River, Vila Nova de Cerveira, Portugal (41º56′20.15″N / 8º44′59.43″W) on May 23, 2022: one designated as neotype (NatMIP-CHAh-0039), and seven others (NatMIP-CHAh-0041).
Diagnosis: Solitary freshwater anthoathecatan brownish polyps, supported posteriorly by flat plate; hypostome hemispherical roof-like, anteriorly with 4–6 tentacles. Ellipsoid stenotele nematocysts, with several size classes (Schuchert 2012). Notes on medusae and reproductive features: No medusae stage. Typical dioecious, remarkably with vegetative reproduction (by buds), smooth to slightly prickly mature eggs, embryotheca; reproductive season in summer or autumn (Schuchert 2010, 2012).
Geographical distribution: Europe (Schuchert 2010, 2012). Other distributions need revision.
Distribution in Portugal: International Minho River, Vila Nova de Cerveira (new record, this study). This is the first taxonomic record of the species for Portugal.
Ecological notes: Freshwater environments (Pallas 1766); found under stones and on the leaves of aquatic plants (Schuchert 2012), such as E. densa (this study).
Order Leptothecata Cornelius, 1992a, b
Family Campanulariidae Johnston, 1836
Genus Gonothyraea Allman, 1864
Gonothyraea loveni (Allman, 1859) (Fig. 2d).
Synonyms: Laomedea loveni Allman, 1859 (original designation);
Calycella parkeri Hilgendorf, 1898; Gonothyraea hyalina Hincks, 1866; Laomedea hyalina (Hincks, 1866); Obelia loveni (Allman, 1859).
Type material: Syntypes: register code F59297 NMV-OZCAM; ZMB 3523 (GBIF 2023).
Type locality: Cramond Island, Scotland (Allman 1859; Cornelius 1982; Calder 2012).
Material examined: Colonies from International Minho River, Caminha, Portugal (41º52′59″N / 8º50′14″W) collected as glass eel fishing bycatch on May 21, 2020: second half hour, depth of 3.3 m (NatMIP-CHLe-0011), third half hour, depth of 3.4 m (NatMIP-CHLe-0012), fourth half hour, depth of 3.9 m (NatMIP-CHLe-0013).
Diagnosis: Colonies orangish and directed upwards, monosiphonic stem, randomly branched, around 30 mm, with annulation posteriorly, and crawling stolon. Alternate bell-shaped hydrothecal cups, with 14 cusps; gonothecae obconical and straight anteriorly, 2–3 gonophores in each projection (Cornelius 1982; Hayward and Ryland 2017). Notes on medusae and reproductive features: Retained reduced medusae stage until release of planulae (Cornelius 1982; Hayward and Ryland 2017).
Geographical distribution: Atlantic Europe; Mediterranean Sea; Atlantic America (Canada to Caribbean Sea, Argentina); New Zealand (Schuchert 2022).
Distribution in Portugal: Azores, depth of 845 m (Rees and White 1966; Cornelius 1982) and International Minho River, Caminha (new record, this study). The present report is the first record of the species for mainland Portugal.
Ecological notes: Usually in oceanic intertidal zone, but some records at depths reaching 200 m (Cornelius 1982) and one possible record at 845 m in Azores Archipelago (Rees and White 1966); associated with plants, such as brown algae (Hayward and Ryland 2017), e.g. Fucus sp. (this study), as well as animals and inert bottom (Cornelius 1982) and salinity range up to 12 (Calder 1976). Conditions on sampling day: temperature: 18 °C; salinity range: 3.8 – 7.6; oxygen: 10.2 mg/l; pH: 7.5; water velocity range: 0.26 – 0.30 m/s.
Genus Obelia Péron & Lesueur, 1810
Obelia dichotoma (Linnaeus, 1758) (Fig. 2e).
Synonyms: Sertularia dichotoma Linnaeus, 1758 (original designation);
Campanularia brasiliensis Meyen, 1834; Campanularia caulini Delle Chiaje, 1841; Campanularia cavolinii Deshayes & Edwards, 1836; Campanularia cheloniae Allman, 1888; Campanularia dichotoma (Linnaeus, 1758); Campanularia maior Meyen, 1834;
Campanularia obtusidens Jäderholm, 1904; Eucope articulata Agassiz, 1865; Eucope parasitica Agassiz, 1865; Gonothyraea integra Fraser, 1940; Laomedea dichotoma (Linnaeus, 1758); Laomedea divaricata McCrady, 1859; Laomedea gracilis Dana, 1846; Laomedea sargassi Broch, 1913; Obelia Adelungi Hartlaub, 1884; Obelia alternata Fraser, 1938; Obelia angulosa Bale, 1888; Obelia arruensis Marktanner-Turneretscher, 1890; Obelia articulata (Agassiz, 1865); Obelia australis von Lendenfeld, 1885; Obelia biserialis Fraser, 1948; Obelia borealis Nutting, 1901; Obelia braziliensis (Meyen, 1834); Obelia cheloniae (Allman, 1888); Obelia chinensis Marktanner-Turneretscher, 1890; Obelia commissuralis McCrady, 1859; Obelia congdoni Hargitt, 1909; Obelia coughtreyi Bale, 1924; Obelia dubia Nutting, 1901; Obelia equilateralis Fraser, 1938; Obelia everta Hargitt, 1927; Obelia fragilis Calkins, 1899; Obelia gracilis Calkins, 1899; Obelia griffini Calkins, 1899; Obelia helgolandica Hartlaub, 1884; Obelia nigrocaulus Hilgendorf, 1898; Obelia nodosa Bale, 1924; Obelia obtusidens (Jäderholm, 1904); Obelia plicata Hincks, 1868; Obelia pygmaea Coughtrey, 1876; Obelia racemosa Fraser, 1941; Obelia rhunicola Billard, 1901; Obelia sinuosa Leloup, 1935; Obelia solowetzkiana Schydlwsky, 1902; Obelia surcularis Calkins, 1899; Obelia tenuis Fraser, 1938; Obelia undotheca Stechow, 1923; Schizocladium ramosum Allman, 1871.
Type material: Holotype: collection code COAST ID 2A3D2545BF0CFFC18CDF0DAB371FF863.mc.12FC9E0EBF0CFFC18CDF0FAA3048FC12 (Humara-Gil and Cruz-Gómez 2018; GBIF 2023). Paratypes: RBCM 976–00403-001, 976–00403-002. Syntypes: MCZ-IZ COEL-396, 397; NMV-OZCAM F58216, F58756, F59284; USNM 42639, 42,647, 43,758, 70,629, 70,840 (GBIF 2023).
Type locality: “Oceano” (Linnaeus 1758), designated as United Kingdom (Schuchert 2022; GBIF 2023), remarkably SW England (Cornelius 1975; Calder 2012; WoRMS Editorial Board 2023).
Material examined: Colonies from International Minho River, Caminha, Portugal (41º52′59″N / 8º50′14″W) collected as glass eel fishing bycatch on May 21, 2020, third half hour, depth of 3.4 m (NatMIP-CHLe-0010), and beam trawl fishing (NatMIP-CHLe-0014).
Diagnosis: Unbranched or branched alternate hydroids with monosiphonic stems, sometimes polysiphonic posteriorly, up to 350 mm. Internodes with several subdivisions, stem emerging at nodes. Hydrotheca bell-like almost wider than longer and around 15 thin posteriorly transversal lines; hydrothecal rim corrugate-shaped. Gonothecae elongated and uniformly widened, anteriorly tapered and with soft collars (Cornelius 1975; Hayward and Ryland 2017). Notes on medusae and reproductive features: Medusa flat-shaped with four simple lips (Gravili et al. 2015); cnidome constituted by atrichous and basitrichous isorhizas, and atrichous anisorhizas (Bouillon and Boero 2000); reproductive season in summer (Cornelius 1995) or throughout the year in the Mediterranean Sea (Gravili et al. 2015).
Geographical distribution: Atlantic Europe; Mediterranean Sea; Azores Archipelago; Red Sea; South Africa; North-West Atlantic to Caribbean Sea; California; Galapagos Islands; Chile; Argentina; Australia and New Zealand (Stechow 1923; Gravili et al. 2015; Hayward and Ryland 2017; Schuchert 2022).
Distribution in Portugal: Azores Archipelago (Rees and White 1966; Schuchert 2022); Berlengas Archipelago (Nobre 1937); International Minho River, Caminha (new record, this study). The present report is the first record of the species for mainland Portugal.
Ecological notes: Intertidal (Gravili et al. 2015) to subtidal zones at depths reaching about 520 m (Ramil and Vervoort 1992); colonies fixed/epibiotic on plants, sponges, gorgonians and other hydrozoans, polychaete tubes, shells, crustaceans, ascidians, bryozoans (Gravili et al. 2015), and algae, e.g. Zonaria tournefortii (Lamouroux, 1805) (Moura 2011), on motionless/inert bottom (Hayward and Ryland 2017). Found on shell surface of brackish bivalve Cerastoderma edule (Linnaeus, 1758) (this study). Conditions on sampling day: temperature: 18 °C; salinity: 3.8; oxygen: 10.2 mg/l; pH: 7.5; water velocity: 0.26 m/s.
Obelia geniculata (Linnaeus, 1758) (Fig. 2f).
Synonyms: Sertularia geniculata Linnaeus, 1758 (original designation);
Campanularia coruscans Schneider, 1898; Campanularia geniculata (Linnaeus, 1758); Eucope alternata Agassiz, 1865; Eucope diaphana Agassiz, 1862; Eucope fusiformis Agassiz, 1865; Eucope polygena Agassiz, 1865; Laomedea geniculata (Linnaeus, 1758); Laomedea lairii Lamouroux, 1816; Monosklera pusilla von Lendenfeld, 1885; Obelia diaphana (Agassiz, 1862); Obelia geniculata var. gaussi Vanhöffen, 1910; Obelia geniculata var. subsessilis Jäderholm, 1904; Obelia gymnopthalma Spagnoli, 1871.
Type material: LINN 1298.19, 1298.20 (The Linnean Society 2023). Syntype: register code F58768 NMV-OZCAM (GBIF 2023).
Type locality: “Oceano” (Linnaeus 1758), designated as Dover, United Kingdom (Cornelius 1975; Calder 2012, 2019a; GBIF 2023).
Material examined: Colonies from International Minho River, La Guardia, Spain (41°52′17.43″N / 8°52′8.01″W) collected as glass eel fishing bycatch on December 30, 2021 (NatMIP-CHLe-0032).
Diagnosis: Ramified upright monosiphonic stems around 50 mm with hydrothecae goblet-shaped and curved internodes with 1–5 posterior segmentations, emerging from sinuous stolons. Oblong gonothecae anteriorly tapering and moulding small central opening (Cornelius 1975, 1995; Hayward and Ryland 2017). Notes on medusae and reproductive features: Enlarged medusae with small or absent velum, which may perform release from March to November (Cornelius 1995) or January to November in Mediterranean waters (Gravili et al. 2015).
Geographical distribution: Wide global distribution, semi-cosmopolitan (Hayward and Ryland 2017), recorded across all of Europe (including International Minho River, La Guardia, Spain, new record, this study), also found in Atlantic, Indo-Pacific, and Arctic Oceans as well as in Oceania -Macquarie Island (Cornelius 1995).
Distribution in Portugal: Algarve continental shelf (Dexter 1992); Sesimbra (Da Cunha 1940); Sines, Tagus River, Peniche, São Pedro de Moel, Douro River mouth (Da Cunha 1944); Leça da Palmeira, Matosinhos (Nobre 1903a).
Ecological notes: On plants, algae, such as Fucus spp. (Vervoort 1993), leaves of kelp (Da Cunha 1940, 1944), such as Laminaria ochroleuca Pylaie 1824 (Moura 2011); on sponges, other hydrozoans, molluscs, ascidians, bryozoans, seahorse Hippocampus guttulatus Cuvier, 1829, and crustaceans (Gravili et al. 2015), e.g. decapod Polybius henslowii Leach, 1820 (Nobre 1903a); found at depths reaching 35 m (Vervoort 1993). Conditions on sampling day: temperature range: 12.7 – 13.5 °C; salinity range: 3.3 – 35; oxygen range: 9.0—10.6 mg/l; pH range: 7.6 – 8.3; water velocity range: 0.19 – 0.29 m/s.
Superfamily Plumularioidea McCrady, 1859
Family Aglaopheniidae Marktanner-Turneretscher, 1890
Genus Aglaophenia Lamouroux, 1812
Aglaophenia pluma (Linnaeus, 1758) (Fig. 3a).

Hydrozoans collected throughout International Minho River Basin: a. Aglaophenia pluma (Linnaeus 1758); b. Aglaophenia tubulifera (Hincks 1861); c. Gymnangium montagui (Billard 1912); d. Nemertesia antennina (Linnaeus 1758); e. Sertularella ellisii (Deshayes & Milne Edwards, 1836); f. Sertularella polyzonias (Linnaeus 1758); g. Abietinaria abietina (Linnaeus 1758). Scales: 250 µm: d; 500 µm: f; 1 mm: a, b, c, e, g
Synonyms: Sertularia pluma Linnaeus, 1758 (original designation);
Aglaophenia cristata (Lamarck, 1816); Aglaophenia pluma pluma (Linnaeus, 1758); Plumularia cristata Lamarck, 1816; Plumularia pluma (Linnaeus, 1758).
Type material: LINN 1298.11, 1298.12, 1298.13 (The Linnean Society 2023).
Type locality: “Oceano” (Linnaeus 1758), designated as United Kingdom: Gannets Rock, Lundy (GBIF 2023).
Material examined: Colonies from International Minho River, Caminha, Portugal, collected as glass eel fishing bycatch, a) March 14, 2021 (41º52′76.62″N / 8º50′41.70″W): fifth half hour, depth of 2.9 m (NatMIP-CHLe-0001); third half hour, depth of 2.5 m (NatMIP-CHLe-0002); b) May 21, 2020 (41º52′59″N / 8º50′14″W): second half hour, depth of 3.3 m (NatMIP-CHLe-0003); third half hour, depth of 3.4 m (NatMIP-CHLe-0004); fourth half hour, depth of 3.9 m (NatMIP-CHLe-0005).
Diagnosis: Brownish to yellow-green colony plume-like directed upward with monosiphonic stem, about 80 mm. Hydrotheca funnel-like, longer than wide, with nine cusps on rim, median globular nematotheca that emerge 1/2 to 1/3 of hydrotheca, and lateral nematotheca overtaking hydrothecal rim somewhat (Svoboda and Cornelius 1991). Corbulae have 5 to 8 transversal ridges (in males) or fused (in females) (Hayward and Ryland 2017). Notes on medusae and reproductive features: With planular stage, hatched into corbula; reproductive season between April and October (Cornelius 1995) or almost the entire year in the Mediterranean Sea (Gravili et al. 2015).
Geographic distribution: Atlantic Europe; Mediterranean Sea; Macaronesia (excluding Madeira Archipelago); African continent; North Atlantic American continent; Caribbean Sea; Japan; Australia (Svoboda and Cornelius 1991; Ramil et al. 1998; Moura 2011; Schuchert 2022).
Distribution in Portugal: Azores Archipelago (Rees and White 1966; Cornelius 1992b; Moura 2011); Madeira Island, Berlengas Archipelago (Nobre 1937; Moura 2011; Moura et al. 2012b); Algarve: Albufeira, Carvoeiro, Alvor, Lagos, Sagres (Moura 2011), Portimão (Da Cunha 1944), Praia da Luz (Svoboda and Cornelius 1991) and continental shelf (Dexter 1992); Vila Nova de Milfontes, Pessegueiro Island, Sesimbra, São Julião da Barra, Cascais (Da Cunha 1940); Porto Covo, Sines, Viana do Castelo (Moura 2011); Setubal, Cruz Quebrada, Oeiras, Cascais, Figueira da Foz (Buarcos), Douro River mouth (Da Cunha 1944); Estoril (Nobre 1903b); Leça da Palmeira, Matosinhos (Nobre 1903a); International Minho River, Caminha (new record, this study).
Ecological notes: Colonies may grow on seaweeds (Gravili et al. 2015), such as Halidrys spp., Cystoseira spp., Sargassum spp. Laminaria spp., on the decapod Maja squinado (Herbst, 1788), as well as on sponges (Da Cunha 1940), rocks (Moura 2011), carbonate concretions, anthozoans and other hydrozoans, shells, crustaceans and bryozoans (Gravili et al. 2015); reaching depths of 20 m, between rock channels in deep pools containing aquatic plants—possibly eelgrasses (Nobre 1903a; Teissier 1965; Svoboda and Cornelius 1991; Hayward and Ryland 2017), and on seagrass Posidonia sp. (Gravili et al. 2015). Conditions on sampling day (March 14): temperature: 13.5 °C; salinity 34.4; oxygen: 9.1 mg/l; pH: 8.2; water velocity: 0.23 m/s; on May 21, temperature: 18 °C; salinity range: 1.1 – 7.6; oxygen: 10.3 mg/l; pH: 7.7; water velocity range: 0.20 – 0.30 m/s.
Aglaophenia tubulifera (Hincks, 1861) (Fig. 3b).
Synonyms: Plumularia tubulifera Hincks, 1861 (original designation);
Aglaophenia filicula Allman, 1883; Plumularia tubulifera Hincks, 1861
Type material: Possibly deposited at Great North Museum: Hancock (Svoboda and Cornelius 1991).
Type locality: SW England: Cornwall (Hincks 1861).
Material examined: Colonies from International Minho River, Caminha, Portugal, collected as glass eel fishing bycatch, a) May 21, 2020 (41º52′59″N / 8º50′14″W): second half hour, depth of 3.3 m (NatMIP-CHLe-0006); b) March 14, 2021 (41º52′76.62″N / 8º50′41.70″W): fifth half hour, depth of 2.9 m (NatMIP-CHLe-0007).
Diagnosis: Colonial structure upwards, with monosiphonic feathered stems to around 60 mm. Hydrotheca gutter-like, anteriorly nine cusps on rim (median one longer than others), medially with nematotheca tube-like extending almost outward to hydrothecal cusps. Corbulae with 8 to 13 transversal ridges (Hayward and Ryland 2017). Notes on medusae and reproductive features: Reproductive season in summer (Cornelius 1995).
Geographical distribution: Eastern North Atlantic, Mediterranean Sea (Svoboda and Cornelius 1991; Hayward and Ryland 2017; Schuchert 2022).
Distribution in Portugal: Azores Archipelago (Rees and White 1966; Cornelius 1992b; Moura 2011); Sesimbra (Svoboda and Cornelius 1991); Berlengas Archipelago (Da Cunha 1944; Moura 2011), Viana do Castelo (Moura et al. 2012b); and International Minho River, Caminha (new record, this study).
Ecological notes: Found on hard bottoms, such as rock (Moura 2011), encrusting bryozoan Smittina spp., seaweed Halidrys spp. (Teissier 1965) and coralline Jania rubens (Linnaeus 1758) (Da Cunha 1944); typical subtidal (Hayward and Ryland 2017), at depths of 8 to 98 m (Rees and White 1966). Conditions on sampling day (March 14): temperature range: 11.3 – 13.5 °C; salinity range: 0.24 – 35.0; oxygen range: 9.1 – 11.1 mg/l; pH range: 7.4 – 8.3; water velocity range: 0.13 – 0.23 m/s. On May 21, temperature: 18.2 °C; salinity: 1.1; oxygen: 10.3 mg/l; pH: 7.3; water velocity: 0.20 m/s.
Genus Gymnangium Hincks, 1874.
Gymnangium montagui (Billard, 1912) (Fig. 3c).
Synonyms: Halicornaria montagui Billard, 1912 (original designation);
Gymnangium pennatulum (Ellis & Solander, 1786).
Type material: Unknown (GBIF 2023). Thus, we propose here the designation of a neotype in the same geographical type locality (European Northeast Atlantic Ocean), because name-bearing type specimen (i.e. holotype, lectotype, syntype or prior neotype) is possibly lost/destroyed (Article 75 and qualifying conditions ( ICZN 1999)): NatMIP-CHLe-0008 from International Minho River, deposited at Natural History Museum of Iberian Peninsula, Vila Nova de Cerveira, Portugal (this study).
Type locality: France: Roscoff (Billard 1912), International Minho River (this study).
Material examined: Colonies from International Minho River, Caminha, Portugal, collected as glass eel fishing bycatch, May 21, 2020 (41º52′59″N / 8º50′14″W): third half hour, depth of 3.4 m (NatMIP-CHLe-0008); fourth half hour, depth of 3.9 m (NatMIP-CHLe-0009).
Diagnosis: Upward yellowish colony feather-like with monosiphonic stem up to 150 mm and alternate ramifications arranged in two rows. Hydrotheca goblet-like with two opposite cusps and with external median tapered nematotheca extending outward beyond edge of hydrotheca, free for most of length. Unclosed gonothecae supported on a stem (Billard 1912; Hayward and Ryland 2017). Notes on medusae and reproductive features: Reproductive season in August (Teissier 1965).
Geographical distribution: Eastern North Atlantic, Mediterranean Sea (Hayward and Ryland 2017; Schuchert 2022).
Distribution in Portugal: Sagres, Porto Covo (Moura 2011); Sines, Berlengas Archipelago, Viana do Castelo (Moura 2011; Moura et al. 2012b); and International Minho River, Caminha (new record, this study).
Ecological notes: Typical subtidal (may be found in partially exposed environment) on hard substrate (Hayward and Ryland 2017), such as rock (Moura 2011), at depths of 60 to 80 m (Teissier 1965). Conditions on sampling day: temperature: 15.4 °C; salinity range: 18.0 – 28.4; oxygen: 9.5.2 mg/l; pH: 8.5; water velocity range: 0.18 – 0.24 m/s.
Family Plumulariidae McCrady, 1859
Genus Nemertesia Lamouroux, 1812
Nemertesia antennina (Linnaeus, 1758) (Fig. 3d).
Synonyms: Sertularia antennina Linnaeus, 1758 (original designation);
Antennularia antennina (Linnaeus, 1758); Antennularia cruciata Pieper, 1881; Antennularia indivisa Lamarck, 1816; Antennularia pentasticha Pieper, 1880; Nemertesia antennina irregularis (Quelch, 1885); Nemertesia irregularis (Quelch, 1885); Nemertesia perrieri var. antennoides (Billard, 1901); Nemertesia pinnata (Nutting, 1900).
Type material: LINN 1298.15 (The Linnean Society 2023).
Type locality: “Oceano” (Linnaeus 1758), designated as United Kingdom (Moura et al. 2012a).
Material examined: Colonies from International Minho River, La Guardia, Spain, collected as glass eel fishing bycatch, December 30, 2021 (41º52′17.43″N / 8º52′8.01″W) (NatMIP-CHLe-0031).
Diagnosis: Erect monosiphonic unbranched stems reaching 350 mm, with two internodes between goblet-shaped hydrothecae. Hydrocaulus coenosarc yellowish-orange (Gravili et al. 2015). Oblong one-side curved gonothecae (Cornelius 1995; Hayward and Ryland 2017). Notes on medusae and reproductive features: Planular dispersion (one per gonotheca), with modified small medusae (Cornelius 1995); reproductive season in September in Mediterranean waters—Tyrrhenian Sea (Stechow 1923).
Geographical distribution: Arctic and Atlantic Oceans, Mediterranean Sea (Stechow 1923; Cornelius 1995; Hayward and Ryland 2017); International Minho River, La Guardia, Spain (new record, this study).
Distribution in Portugal: Azores (Rees and White 1966; Moura 2011) and Madeira archipelagos (Cornelius 1995); Sagres, Porto Covo, Sines, Berlengas Archipelago (Moura 2011); Sado River estuary (Dexter 1992); Setubal (Da Cunha 1940, 1944); Estoril (Nobre 1903b); Peniche (Nobre 1937); Porto municipality (Da Cunha 1944); Póvoa de Varzim (Nobre 1903a); Viana do Castelo (Moura 2011).
Ecological notes: Found from offshore to the sublittoral zone (Hayward and Ryland 2017) reaching depths of 1500 m (Gravili et al. 2015). Sampled with fishing nets denominated “rasqueiras” (Nobre 1903a). Colonies grow on hard bottoms, e.g. rock (Moura 2011), corals, and shells (Gravili et al. 2015), as well as on sandy beaches (Hayward and Ryland 2017), at depths reaching 80 m (Vervoort 1993). Conditions on sampling day: temperature range: 12.7 – 13.5 °C; salinity range: 3.3 – 35; oxygen range: 9.0—10.6 mg/l; pH range: 7.6 – 8.3; water velocity range: 0.19 – 0.29 m/s.
Superfamily Sertularioidea Lamouroux, 1812
Family Sertularellidae Maronna, Miranda, Peña Cantero, Barbeitos, Marques, 2016
Genus Sertularella Gray, 1848
Sertularella ellisii (Deshayes & Milne Edwards, 1836) (Fig. 3e).
Synonyms: Sertularia ellisii Deshayes & Milne-Edwards, 1836 (original designation);
Sertularella polyzonias var. ellisi.
Type material: Unknown.
Type locality: European seas (Deshayes and Milne-Edwards 1836), designated as north coast of Kent, England (Vervoort 1993).
Material examined: Colonies from International Minho River, Caminha, Portugal, collected as glass eel fishing bycatch, May 21, 2020): second half hour, depth of 3.3 m (41º52′59.00″N / 8º50′14.00″W) (NatMIP-CHLe-0035).
Diagnosis: Upright monosiphonic stems, ramified softly, reaching to around 35 mm; hydrothecae bottle-shaped with four cusps anteriorly. Oblong segmented gonothecae with tapered opening (Ramil et al. 1992; Hayward and Ryland 2017). Notes on medusae and reproductive features: With planular stage, incubated in external acrocyst (Cornelius 1995); reproductive season from February to September (Ramil et al. 1992) or January to November in Mediterranean Sea (Gravili et al. 2015).
Geographical distribution: Atlantic Europe and Mediterranean Sea (Ramil et al. 1992; Moura 2011; Gravili et al. 2015; Hayward and Ryland 2017).
Distribution in Portugal: Azores and Madeira Archipelagos (Medel and Vervoort 1998); Sagres, Cascais, Berlengas Archipelago (Moura 2011); International Minho River, Caminha (new record, this study).
Ecological notes: Colonies grow on sandy and muddy environments (Vervoort 1993), rocky bottom (Moura 2011), red algae Corallina spp., brown algae Cystoseira spp., seagrass Posidonia spp., ascidians, sponges, and other hydrozoans (Gravili et al. 2015) from intertidal to sublittoral habitats (Hayward and Ryland 2017) at around to 90 m in depth (Gravili et al. 2015). Conditions on sampling day: temperature: 15.4 °C; salinity: 9.1; oxygen: 10.1 mg/l; pH: 7.9; water velocity: 0.24 m/s.
Remarks: Despite the current validity of this species (WoRMS Editorial Board 2023), a review of the phylogenetic systematics is needed due to its nomenclatural instability (Cornelius 1979, 1995).
Sertularella polyzonias (Linnaeus, 1758) (Fig. 3f).
Synonyms: Sertularia polyzonias Linnaeus, 1758 (original designation);
Sertularella falsa Millard, 1957.
Type material: LINN 1298.21, LINN 1298.22, LINN 1298.23, LINN 1298.24 (The Linnean Society 2023). Paratype (GBIF Taxon interpretation): Deposited at NHMUK, register code 1957.4.26.42, labelled as Sertularella falsa Millard, 1957 (GBIF 2023).
Type locality: “Oceano” (Linnaeus 1758), north coast of Kent, England (Cornelius 1979; Calder 2012).
Material examined: Colonies from International Minho River, Caminha, Portugal, collected as glass eel fishing bycatch, March 14, 2021): third half hour, depth of 2.5 m (41º52′76.62″N / 8º50′41.70″W) (NatMIP-CHLe-0029).
Diagnosis: Yellowish branched colonies directed upright with monosiphonic stem around 50 mm. Smooth to softly wrinkled hydrothecae anteriorly with 4 cusps; elliptical crinkled gonothecae (Cornelius 1995; Hayward and Ryland 2017). Notes on medusae and reproductive features: With planktonic larval stage, brooding in acrocyst; reproductive season in summer (Cornelius 1979, 1995) or throughout the entire year in Mediterranean waters (Gravili et al. 2015).
Geographical distribution: Atlantic and Arctic Oceans, Mediterranean Sea, Red Sea (Cornelius 1995; Hayward and Ryland 2017; WoRMS Editorial Board 2023).
Distribution in Portugal: Azores Archipelago (Rees and White 1966; Cornelius 1992b; Moura 2011); Algarve: Olhão (Nobre 1937), Faro (Culatra Island) (Da Cunha 1944), continental shelf (Dexter 1992); Sines, Sesimbra, Porto municipality (Da Cunha 1944); Berlengas Archipelago (Moura 2011); Póvoa de Varzim (Nobre 1903a); Viana do Castelo (Moura 2011); and International Minho River, Caminha (new record, this study).
Ecological notes: This species may be found along the coast collected in fishing nets denominated “rasqueiras” (Nobre 1903a) from the intertidal to subtidal zone at depths reaching 300 m (Hayward and Ryland 2017). Colonies grow on algae, seagrasses, such as Posidonia oceanica (Linnaeus 1767) and Zostera spp., on other hydrozoans, and calcareous substrata, including polychaete tubes and dead gorgonians (Gravili et al. 2015). Conditions on sampling day: temperature: 13.4 °C; salinity: 32.5; oxygen: 9.2 mg/l; pH: 8.2; water velocity: 0.27 m/s.
Family Sertulariidae Lamouroux, 1812
Genus Abietinaria Kirchenpauer, 1884
Abietinaria abietina (Linnaeus, 1758) (Fig. 3g).
Synonym: Sertularia abietina Linnaeus, 1758 (original designation).
Type material: LINN 1298.4 (The Linnean Society 2023).
Type locality: “Oceano” (Linnaeus 1758), designated as south coast of England (Cornelius 1979; Calder 2012).
Material examined: Colonies from International Minho River, La Guardia, Spain, collected as glass eel fishing bycatch, December 30, 2021 (41º52′17.43″N / 8º52′8.01″W) (NatMIP-CHLe-0030).
Diagnosis: Erect randomly branched monosiphonic stems up to 300 mm; hydrothecae bottle-shaped (some anteriorly pink to reddish, this study) with anterior opening curved dorsally. Lengthened-oblong gonothecae tapered posteriorly with anterior end (Cornelius 1979, 1995; Hayward and Ryland 2017). Notes on medusae and reproductive features: Embryos emerging in external acrocyst; reproductive season in winter and spring (Cornelius 1979, 1995; Hayward and Ryland 2017).
Geographical distribution: Baltic Sea; North Atlantic Ocean, Mediterranean Sea, North Pacific Ocean, Japan, Madagascar (Cornelius 1979, 1995; Hayward and Ryland 2017); International Minho River, La Guardia, Spain (new record, this study).
Distribution in Portugal: Madeira Archipelago (Cornelius 1995); Berlengas Archipelago (Nobre 1937), Porto municipality (Da Cunha 1944); Viana do Castelo (Moura 2011); Caminha (Moledo do Minho) (Nobre 1903a).
Ecological notes: On algae (Nobre 1903a), pebbles, shells, and sandy bottoms, at depths reaching 10 m (Cornelius 1979, 1995; Hayward and Ryland 2017). Conditions on sampling day: temperature range: 12.7 – 13.5 °C; salinity range: 3.3 – 35; oxygen range: 9.0—10.6 mg/l; pH range: 7.6 – 8.3; water velocity range: 0.19 – 0.29 m/s.
Genus Amphisbetia Agassiz, 1862
Amphisbetia distans (Lamouroux, 1816) (Fig. 4a).

Hydrozoans collected throughout International Minho River Basin: a. Amphisbetia distans (Lamouroux 1816); b. Amphisbetia operculata (Linnaeus 1758); c. Diphasia fallax (Johnston, 1847); d. Dynamena pumila (Linnaeus 1758); e. Sertularia cupressina Linnaeus 1758; f. Thuiaria articulata (Pallas 1766). Scales: 500 µm: a, c, d; 1 mm: b, e, f
Synonyms: Dynamena distans Lamouroux, 1816 (original designation);
Dynamena secunda Heller, 1868; Sertularia distans (Lamouroux, 1816); Sertularia distans var. gracilis (Lamouroux, 1816); Sertularia erasmoi Vannucci, 1946; Sertularia gracilis Hassal, 1848; Sertularia lamourousii Deshayes & Milne Edwards, 1836; Sertularia stookeyi Nutting, 1904; Tridentata distans (Lamouroux, 1816).
Type material: Possible holotype (but no site/museum designation): taxon ID 9E4CE23AFF81F10EFF0360DBFA7429F1.taxon (GBIF 2023). Syntypes: USNM 19710, 19,711; NMV-OZCAM F60330 (GBIF 2023).
Type locality: Atlantic Ocean (Lamouroux 1816), possibly in northern zone.
Material examined: Colonies from International Minho River, Caminha, Portugal, collected as glass eel fishing bycatch, May 21, 2020 (41º52′59″N / 8º50′14″W): first half hour, depth of 3.3 m (NatMIP-CHLe-0019); second half hour, depth of 3.3 m depth (NatMIP-CHLe-0020); third half hour, depth of 3.4 m (NatMIP-CHLe-0021); fourth half hour, depth of 3.9 m (NatMIP-CHLe-0022).
Diagnosis: Uniformly brownish erect colonies, approximately 30 mm, ramified little, with hydrothecal cups symmetrically arranged about 30% anteriorly directed outward, tapering posterior end with 2–3 teeth. Internode evenly of same width throughout colony. Gonothecae elliptical with anterior opening. Notes on medusae and reproductive features: With planular stage, possibly hatched in acrocyst; reproductive season spring to summer (Cornelius 1995).
Geographical distribution: British Isles, Mediterranean Sea (Ramil and Vervoort 1992; Hayward and Ryland 2017), France: Roscoff (Billard 1912), Portugal (Nobre 1903a; Cornelius 1992b; Moura 2011), North-West Atlantic (Calder 2019a; Schuchert 2022).
Distribution in Portugal: Azores (Rees and White 1966; Cornelius 1992b) and Madeira Archipelagos (Moura 2011); Sagres, Aljezur, Vila Nova de Milfontes, Porto Covo, Sines, Cascais, Ericeira, Berlengas Archipelago (Moura 2011); Sesimbra (Da Cunha 1944); Douro River mouth (Leixões Port) (Nobre 1903a; Da Cunha 1944); Viana do Castelo (Moura 2011); and International Minho River, Caminha (new record, this study).
Ecological notes: Usually found in intertidal to subtidal zone up to 60 m depth (Calder 2019a), and possibly until 826 m (Ramil and Vervoort 1992), on rock (Moura 2011), on seaweed surfaces (Nobre 1903a), e.g. Sargassum spp. (Calder 2019a), and associated with bryozoans, other hydrozoan species, and eventually other organisms (Hayward and Ryland 2017). Conditions on sampling day: temperature: 18 °C; salinity range: 1.1 – 7.6; oxygen: 10.3 mg/l; pH: 7.7; water velocity range: 0.20 – 0.30 m/s.
Amphisbetia otalizing (Linnaeus, 1758) (Fig. 4b).
Synonyms: Sertularia otalizing Linnaeus, 1758 (original designation);
Amphisbetia pulchella (D’Orbigny, 1846); Dynamena otalizing (Linnaeus, 1758); Dynamena pulchella d’Orbigny, 1842; Sertularia aperta Allman, 1885.
Type material: NHMUK 1927.1.1.2 / ecatalogue: 658,656 (GBIF 2023).
Type locality: “Oceano” (Linnaeus 1758), designated as South Africa: Cape of Good Hope (GBIF 2023).
Material examined: Colonies from International Minho River, Caminha, Portugal, collected as glass eel fishing bycatch, May 21, 2020 (41º52′59″N / 8º50′14″W): fourth half hour, depth of 3.9 m (NatMIP-CHLe-0023).
Diagnosis: Colonies growing upright, reaching 100 mm, at times 160 mm (Da Cunha 1940), sprawling growth, hydrotheca cylindrical-shaped (Cornelius 1995), arranged suboppositely, with two asymmetrical cusps (Hayward and Ryland 2017). Smooth gonothecae globular-like with opening at anterior edge (Hayward and Ryland 2017). Colour yellowish with reddish ribs (above each hydrothecal pair). Notes on medusae and reproductive features: Individuals are released at sunrise (Teissier 1922), small ones mature into gonotheca (Hayward and Ryland 2017), reproductive season spring to summer (Cornelius 1995).
Geographical distribution: Europe (Lamouroux 1816; Schuchert 2022): British Isles (Cornelius 1995), France (Billard 1912), Portugal (Da Cunha 1940; Cornelius 1992b), Mediterranean Sea (Hayward and Ryland 2017); Java Island, Africa (Cornelius 1995), Argentina (Genzano 1990), Chile (Cornelius 1995); New Zealand (Cairns et al. 2009).
Distribution in Portugal: Azores Archipelago (Cornelius 1992b; Moura 2011); Sagres (Nobre 1937), Aljezur, Porto Covo (Moura 2011); Pessegueiro Island, Berlengas Archipelago (Da Cunha 1940); Sines, Setubal, Douro River mouth (Da Cunha 1944); Viana do Castelo (Moura 2011); and International Minho River, Caminha (new record, this study).
Ecological notes: Associated to kelp holdfasts (Billard 1912; Cornelius 1995), on cirriped Balanus spp. (Da Cunha 1940) and on rock (Moura 2011). Conditions on sampling day: temperature: 18.0 °C; salinity 7.6; oxygen: 10.3 mg/l; pH: 7.9; water velocity: 0.3 m/s.
Genus Diphasia Agassiz, 1862
Diphasia fallax (Johnston, 1847) (Fig. 4c).
Synonyms: Sertularia fallax Johnston, 1847 (original designation);
Diphasia wandeli Levinsen, 1893; Nigellastrum fallax (Johnston, 1847).
Type material: NHMD-98287 (GBIF 2023). Possibly syntypes: BM(NH)-NHMUK 1847.9.22.24a; 1847.9.22.29; 1847.9.22.30; 1847.9.22.31 (Cornelius 1979).
Type locality: North Sea, northeast coast of Britain (Cornelius 1979; Calder 2012; WoRMS Editorial Board 2023).
Material examined: Colonies from International Minho River, Caminha, Portugal, collected as glass eel fishing bycatch, March 14, 2021: fifth half hour, depth of 2.5 m (41º52′76.62″N / 8º50′41.70″W) (NatMIP-CHLe-0036).
Diagnosis: Upright-branched monosiphonic stems up to 100 mm, with tubular subadnate hydrothecae arranged in pairs side by side. Gonothecae inverted bottom-shaped anteriorly 4-horned (spines) (Cornelius 1979, 1995; Hayward and Ryland 2017). Notes on medusae and reproductive features: With planular stage, hatched in acrocyst; reproductive season between May and July (Cornelius 1979, 1995; Hayward and Ryland 2017).
Geographical distribution: North European and American Atlantic (USA) to Arctic Oceans (Cornelius 1995; Hayward and Ryland 2017; Gil and Ramil 2017). The present report is the first record of the species for Southern Europe (new record, this study).
Distribution in Portugal: International Minho River, Caminha (new record, this study).
Ecological notes: In middle sublittoral waters to depths of 250 m (Hayward and Ryland 2017); epizoic on hydrozoans Aglaophenopsis spp. And Thuiaria spp. (Cornelius 1995). Conditions on sampling day: temperature: 13.5 °C; salinity 34.4; oxygen: 9.1 mg/l; pH: 8.3; water velocity: 0.23 m/s.
Genus Dynamena Lamouroux, 1812
Dynamena pumila (Linnaeus, 1758) (Fig. 4d).
Synonym: Sertularia pumila Linnaeus, 1758 (original designation).
Type material: LINN 1298.1, 1298.2 (The Linnean Society 2023).
Type locality: “Oceano” (Linnaeus 1758), designated as Rottingdean, Sussex, England (Cornelius 1979; Calder 2012; GBIF 2023).
Material examined: Colonies from International Minho River, Caminha, Portugal (41º52′59″N / 8º50′14″W), collected as glass eel fishing bycatch on May 21, 2020: first half hour, depth of 3.3 m (NatMIP-CHLe-0015), second half hour, depth of 3.3 m (NatMIP-CHLe-0016), third half hour, depth of 3.4 m (NatMIP-CHLe-0017), fourth half hour, depth of 3.9 m (NatMIP-CHLe-0018).
Diagnosis: Erect unramified monosiphonic stems around 30 mm, with tubular subadnate hydrothecae arranged in pairs side by side. Oblong gonothecae softly wrinkled with enlarged anterior opening (Cornelius 1995; Hayward and Ryland 2017). Notes on medusae and reproductive features: With planular stage hatched in acrocyst; reproductive season spring to summer (Cornelius 1995; Hayward and Ryland 2017).
Geographical distribution: Arctic Ocean to North European Atlantic (Cornelius 1995; Calder 2012; Hayward and Ryland 2017).
Distribution in Portugal: Berlengas Archipelago (Da Cunha 1944); Ílhavo (Barra Beach) (Moura 2011); Douro River mouth (Nobre 1903a); International Minho River, Caminha (new record, this study).
Ecological notes: From intertidal to upper sublittoral zones (Cornelius 1995), in marine and brackish environments (Hayward and Ryland 2017) on seaweed (Moura 2011), such as Ascophyllum nodosum (Linnaeus, 1753) and Fucus spp. (Cornelius 1979), rocky shore (Hayward and Ryland 2017), encrusting algae and sponges on rocks (Nobre 1903a). Conditions on sampling day: temperature: 18 °C; salinity range: 1.1 – 7.6; oxygen: 10.3 mg/l; pH: 7.7; water velocity range: 0.20 – 0.30 m/s.
Genus Sertularia Linnaeus 1758
Sertularia cupressina Linnaeus, 1758 (Fig. 4e).
Synonym: Thuiaria cupressina Linnaeus, 1758
Type material: LINN 1298.5, 1298.6 (Cornelius 1979; The Linnean Society 2023).
Type locality: “Oceano” (Linnaeus 1758), designated as India (The Linnean Society 2023).
Material examined: Colonies from International Minho River, Portugal: Caminha, collected as glass eel fishing bycatch, May 21, 2020 (41º52′59″N / 8º50′14″W): first half hour, depth of 3.3 m (NatMIP-CHLe-0024); second half hour, depth of 3.3 m (NatMIP-CHLe-0025); third half hour, depth of 3.4 m (NatMIP-CHLe-0026); fourth half hour, depth of 3.9 m (NatMIP-CHLe-0027); Cristêlo, through Van Veen dredging, September 24, 2020 (42º00′58.20″N / 8º39′29.09″W) (NatMIP-CHLe-0028).
Diagnosis: Tuft-like spread helical monosiphonic branched stems around 600 mm, known as “sea cypress” (Cornelius 1995; Hayward and Ryland 2017), hydrothecae quasi-adnate arranged semi-oppositely, posterior section enlarged and tapering anteriorly (Cornelius 1979). Gonothecae narrowed putter-shaped, distally with ring-shaped openings and lateral spines (Cornelius 1995). Notes on medusae and reproductive features: With planular stage, growing in acrocyst (Hayward and Ryland 2017); reproductive season from March to November (Cornelius 1979, 1995).
Geographical distribution: Arctic Ocean to North European and American Atlantic, Mediterranean Sea, North Pacific Ocean (Cornelius 1979, 1995; Hayward and Ryland 2017; WoRMS Editorial Board 2023).
Distribution in Portugal: International Minho River: municipalities of Caminha and Valença (Cristêlo) (new record, this study). This is the first taxonomic record of the species for Portugal.
Ecological notes: Found in sandy and rocky marine environments at depths to 100 m (Cornelius 1979, 1995; Hayward and Ryland 2017) and on river sediment (this study). Conditions on sampling day: temperature: 18 °C; salinity range: 1.1 – 7.6; oxygen: 10.3 mg/l; pH: 7.7; water velocity range: 0.20 – 0.30 m/s.
Genus Thuiaria Fleming, 1828
Thuiaria articulata (Pallas, 1766) (Fig. 4f).
Synonyms: Sertularia articulata Pallas, 1766 (original designation);
Dymella articulata (Pallas, 1766); Salacia articulata (Pallas, 1766); Sertularia lonchitis Ellis & Solander, 1786; Thuiaria barentsi Naumov, 1960; Thuiaria kolaënsis Jäderholm, 1909; Thuiaria lichenastrum (Linnaeus, 1758); Thuiaria lonchitis (Ellis & Solander, 1786).
Type material: Neotype: register code 1893.2.28.13, labelled as Sertularia articulata Pallas, 1766 (Cornelius 1979). Paratype: register code NHMUK 1962.10.10.21 / ecatalogue: 668,969, labelled as Thuiaria barentsi Naumov, 1960 (GBIF 2023).
Type locality: Lancashire, England (Cornelius 1979). White Sea, Russia (based on paratype) (GBIF 2023).
Material examined: Colonies from International Minho River, La Guardia, Spain, collected as glass eel fishing bycatch, December 30, 2021 (41º52′17.43″N / 8º52′8.01″W) (NatMIP-CHLe-0033).
Diagnosis: Upright brownish branched stems up to 250 mm, alternate hydrothecae goblet-like and anterior tapered end. Gonothecae slightly ellipsoid with opening ring-shaped (Cornelius 1979, 1995; Hayward and Ryland 2017). Notes on medusae and reproductive features: With planular stage, hatched in acrocyst (Hayward and Ryland 2017); reproductive season from March to May (Cornelius 1979, 1995).
Geographical distribution: Arctic Ocean to United Kingdom; International Minho River, La Guardia, Spain (new record, this study).
Distribution in Portugal: Viana do Castelo (Moura 2011).
Ecological notes: Found on hard bottom, e.g. rock (Moura 2011) and shells (Cornelius 1995) in sublittoral zone from depths of 18 to 300 m (Hayward and Ryland 2017). Conditions on sampling day: temperature range: 12.7 – 13.5 °C; salinity range: 3.3 – 35; oxygen range: 9.0 – 10.6 mg/l; pH range: 7.6 – 8.3; water velocity range: 0.19 – 0.29 m/s.
Discussion
The first taxonomic/checklist surveys in Portugal were performed by Dr. Augusto Nobre and provided a species list for northern (Nobre 1903a) and southern Portugal (Nobre 1903b) comprising 25 species belonging to the Class Hydrozoa. The species were sampled on several substrates (including rocky and sandy beaches), such as algae, encrusting sponges, bivalves, gastropods, crustaceans, rocky shores, harbour buoys, and fishing traps as well as trawl fishing nets, cast net, and “rasqueira” nets (Nobre 1903a, b). Between the 1930s and 1950s, Dr. Augusto Nobre and Dr. Alberto da Cunha developed more checklists (including descriptions, drawings, and identification keys) of the hydrozoan species identified along the coast of Portugal, totalizing 50 species (Nobre 1937; Da Cunha 1940, 1944; Moura 2011; TriploV 2022; Universidade do Porto 2022).
In 1966, Rees and White compiled all data on hydrozoans from the Azores Archipelago collected in previous expeditions, e.g. Challenger, Imperial College, Queen Mary College, Prince of Monaco, including new records and resulting in a total of 71 species (Rees and White 1966). Paul Cornelius then contributed to the Azorean fauna by adding seven new records of hydrozoans and suggesting that many hydrozoan species are dispersed by rafting across aquatic environments (Cornelius 1992b).
In the early 1990s, Deborah Dexter compiled data from previous surveys on invertebrates collected on soft substrates along the Portuguese coast, reporting 15 hydrozoan species for the Algarve continental shelf, Sado River, Tagus River, and Figueira da Foz (Dexter 1992). Carlos Moura also compiled data on Portuguese hydrozoans, providing a list of species and localities collected along the coast, also including the Azores and Madeira Archipelagos (Moura 2011).
The species in the present study were mainly collected as bycatch of glass eel fishing activities during the new moon using anchored stow nets (passive gear) (Weber 1986; Antunes and Weber 1996). This is in agreement with the rafting dispersal hypothesis put forth by Cornelius (see above) for most species caught in the present study and with our observations, remarkably for species found in the tidal marine and brackish zones, with the direct influence of seawater.
With the present work, we have added four new records for Portugal and one for southern Europe, demonstrating the strong potential for finding more hydrozoan species with an increased sampling effort. Considering the lack of some type material for many cnidarian species (and also many invertebrates), this study offers three neotypes, filling a gap of resulting from the absence/loss of name-bearing types commonly detected for species described in the 18th and 19th Centuries.
Conclusions
This study constitutes the first hydrozoan taxonomic survey of the International Minho River, with a total of 19 species, four of which are new records for Portugal, six are new records for mainland Portugal, and one is a new record for Southern Europe (Diphasia fallax).
Most species were collected as glass eel fishing bycatch; one by beam trawl (Obelia dichotoma), one using Van Veen dredging (Sertularia cupressina); one collected by hand on wood fragments (Cordylophora caspia); and two sampled by hand on the leaves of Egeria densa (Hydra viridissima and Hydra oligactis). The International Minho River is a potential site for the dispersal and occurrence of numerous hydrozoan species, mainly through an analysis of glass eel fishing bycatch.
Considering the representativeness of these important bioindicators, this study can contribute to future studies on environmental impacts, coastal management, and conservation in the Minho River, with a possible influence on surveys in nearby coastal areas.
Change history
01 September 2023
A Correction to this paper has been published: https://doi.org/10.1007/s11852-023-00972-w
References
Allman GJ (1859) Notes on the hydroid zoophytes. Ann Mag Nat Hist 4:137–144
Antunes C, Weber M (1996) The glass eel fishery and the by-catch in the Rio Minho after one decade (1981–1982 and 1991–1992). Arch Polish Fish 4:131–139
Billard A (1912) Hydroïdes de Roscoff. Arch Zool Expérimentale Générale 51:459–478
Bouillon J, Boero F (2000) Phylogeny and classification of Hydroidomedusae: synopsis of the families and genera of the Hydromedusae of the world, with a list of the worldwide species. Thalassia Salentina 24:47–296. https://doi.org/10.1285/i15910725v24p47
Bouillon J, Gravili C, Pagès F et al (2006) An introduction to Hydrozoa. Mémoires du Muséum national d’Histoire naturelle, Tome 194, Paris
Cairns SD, Gershwin L-A, Brook FJ et al (2009) Phylum Cnidaria: corals, medusae, hydroids, myxozoans. In: Gordon DP (ed) New Zealand inventory of biodiversity: 1. Kingdom Animalia: Radiata, Lophotrochozoa, Deuterostomia. Smithsonian Libraries, Washington, pp 59–101
Calder DR (2019a) On a collection of hydroids (Cnidaria, Hydrozoa) from the southwest coast of Florida, USA. Zootaxa 4689:1–141. https://doi.org/10.11646/zootaxa.4689.1.1
Calder DR (2019b) On a collection of hydroids (Cnidaria, Hydrozoa) from the southwest coast of Florida, USA. In: GBIF.org/Plazi.org Taxon. Treat. database. https://www.gbif.org/dataset/a63613b4-e611-42e8-89cd-2d3aae126759. Accessed 31 May 2022
Calder DR (2012) On a collection of hydroids (Cnidaria, Hydrozoa, Hydroidolina) from the west coast of Sweden, with a checklist of species from the region. Zootaxa 3171:1–77. https://doi.org/10.11646/zootaxa.3171.1.1
Calder DR (1976) The zonation of hydroids along salinity gradients in South Carolina estuaries. In: Mackie GO (ed) Third International Conference on Coelenterate Biology. Plenum Publishing Corporation, New York, pp 165–174
Conde A, Domínguez J, Novais JM, Ramil F (2013) First record of Cordylophora caspia (Hydrozoa: Cnidaria) in the Tagus estuary, central Portugal. Mar Biodivers Rec 6:e114(1–6). https://doi.org/10.1017/S1755267213000833
Cornelius PFS (1992a) Medusa loss in leptolid Hydrozoa (Cnidaria), hydroid rafting, and abbreviated life-cycles among their remote-island faunae: an interim review. Sci Mar 56:245–261
Cornelius PFS (1995) North-West European thecate hydroids and their medusae, Part 2: Sertulariidae to Campanulariidae: keys and notes for identification of the species. In: Barnes RSK, Crothers JH (eds) Synopses of the British Fauna (New Series), no. 50. Field Studies Council, Linnean Society of London, Estuarine and Coastal Sciences Association, Shrewsbury, p vii+416
Cornelius PFS (1979) A revision of the species of Sertulariidae (Coelenterata : Hydroida) recorded from Britain and nearby seas. Bull Br Museum Nat Hist 34:243–321
Cornelius PFS (1982) Hydroids and medusae of the family Companulariidae recorded from the eastern North Atlantic, with a world synopsis of genera. Bull Br Museum Nat Hist 42:37–148
Cornelius PFS (1975) The hydroid species of Obelia (Coelenterata, Hydrozoa: Campanulariidae), with notes on the medusa stage. Bull Br Museum Nat Hist Zool 28:249–293. https://doi.org/10.5962/p.314143
Cornelius PFS (1992b) The Azores hydroid fauna and its origin, with discussion of rafting and medusa suppression. Arquipélago - Life Earth Sci 10:75–99
Correia MJ, Costa JL, Chainho P et al (2012) Inter-annual variations of macrobenthic communities over three decades in a land-locked coastal lagoon (Santo André, SW Portugal). Estuar Coast Shelf Sci 110:168–175. https://doi.org/10.1016/j.ecss.2012.04.028
Da Cunha AX (1940) Contribuição para o estudo dos hidropólipos das costas de Portugal (Colecção do Museu Bocage). Arq Do Mus Bocage 11:105–120
Da Cunha AX (1944) Hidropólipos das costas de Portugal. Memórias e Estud Do Mus Zoológico Da Univ Coimbra 161:1–101
Daly M, Brugler MR, Cartwright P, et al (2007) The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. In: Zhang, Z.-Q. & Shear, W.A. (Eds) (2007) Linnaeus Tercentenary: Progress in Invertebrate Taxonomy. Zootaxa, 1668, 1–766. Zootaxa 1668:127–182. https://doi.org/10.11646/zootaxa.1668.1.11
Deshayes GP, Milne-Edwards H (1836) Histoire Naturelle des Animaux sans Vertèbres, présentant les caractères généraux et particuliers de ces animaux, leur distribution, leurs classes, leurs familles, leurs genres, et la citation des principales espèces qui s’y rapportent, par J. B. P. A. de, 2nd edn. J.B. Baillière, Libraire, Paris
Dexter DM (1992) Soft bottom invertebrates of the Portuguese benthos. Bol Do Inst Nac Investig Das Pescas 17:61–88
Encyclopædia Britannica (2023) Hydroid: cnidarian class. https://www.britannica.com/animal/Hydrozoa. Accessed 7 Jan 2023
GBIF (2023) GBIF - Occurrences. In: Glob. Biodivers. Inf. Facil. https://www.gbif.org/occurrence/search/. Accessed 7 Jan 2023
Genzano GN (1990) Hydropolips (Cnidaria) of Mar del Plata, Argentina. Nerítica-Revista Do Cent Biol Mar Da Univ Fed Do Paraná 5:35–54. https://doi.org/10.5380/rn.v5i1.85005
Gil M, Ramil F (2017) The genus Diphasia L. Agassiz, 1862 (Cnidaria, Hydrozoa) in Northwest Africa. Zootaxa 4363:301–349. https://doi.org/10.11646/zootaxa.4363.3.1
Gravili C, Vito D, Camillo CG, et al (2015) The non-Siphonophoran Hydrozoa (Cnidaria) of Salento, Italy with notes on their life-cycles: an illustrated guide. Zootaxa 3908:1–187. https://doi.org/10.11646/zootaxa.3908.1.1
Hamada M, Satoh N, Khalturin K (2020) A reference genome from the symbiotic hydrozoan, Hydra viridissima. G3 Genes|Genomes|Genetics 10:3883–3895. https://doi.org/10.1534/g3.120.401411
Hammond G (2009) Hydrozoa. In: Anim. Divers. Web. https://animaldiversity.org/accounts/Hydrozoa/. Accessed 20 Sep 2021
Hayward PJ, Ryland JS (2017) Handbook of the marine fauna of North-West Europe, 2nd edn. Oxford University Press, New York
Hincks T (1861) XVI.—A catalogue of the zoophytes of South Devon and South Cornwall. Ann Mag Nat Hist Zool Bot Geol 8(ser.3):152–161
Humara-Gil KJ, Cruz-Gómez C (2018) New records of benthic hydroids (Cnidaria: Hydrozoa) from the coast of Oaxaca, Mexico. Zootaxa 4455:454–470. https://doi.org/10.11646/zootaxa.4455.3.3
International Commission on Zoological Nomenclature, Ride WDL, Cogger HG, et al (1999) ICZN - International Code of Zoological Nomenclature. In: Int. Trust Zool. Nomencl. https://code.iczn.org/. Accessed 8 Jan 2022
Lamouroux JVF (1816) Histoire des polypiers coralligènes flexibles, vulgairement nommés zoophytes. F. Poisson, Caen
Linnaeus C (1767) Systema naturae per regna tria naturae: secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tom. 1. Pars. 2, 12. 1. Laurentii Salvii, Holmiae [Stockholm]
Linnaeus C (1758) Systema naturae per regna tria naturae :secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis, Editio dec. Impensis Direct. Laurentii Salvii, Holmiæ [Stockholm]
Medel MD, Vervoort W (1998) Atlantic Thyroscyphidae and Sertulariidae (Hydrozoa, Cnidaria) collected during the CANCAP and Mauritania-II expeditions of the National Museum of Natural History, Leiden, The Netherlands: CANCAP-project. Contributions, no. 120. Zool Verh 320:1–85
Moura CFJ (2011) Systematics and evolution of coastal and deep-water Hydrozoa from the NE Atlantic. University of Aveiro, Aveiro, Portugal. PhD Thesis. https://hdl.handle.net/10773/7570
Moura CJ, Cunha MR, Porteiro FM et al (2012a) Evolution of Nemertesia hydroids (Cnidaria: Hydrozoa, Plumulariidae) from the shallow and deep waters of the NE Atlantic and western Mediterranean. Zool Scr 41:79–96. https://doi.org/10.1111/j.1463-6409.2011.00503.x
Moura CJ, Cunha MR, Porteiro FM, Rogers AD (2012b) A molecular phylogenetic appraisal of the systematics of the Aglaopheniidae (Cnidaria: Hydrozoa, Leptothecata) from the north-east Atlantic and west Mediterranean. Zool J Linn Soc 164:717–727. https://doi.org/10.1111/j.1096-3642.2011.00784.x
Nobre A (1903a) Subsidios para o estudo da fauna marinha do norte de Portugal. Ann Sci Naturaes Rev Hist Nat Agric Piscic e Pescas Maritimas 8:37–94
Nobre A (1903b) Subsídios para o estudo da fauna marinha do sul de Portugal. Ann Sci Naturaes Rev Hist Nat Agric Piscic e Pescas Maritimas 8:153–160
Nobre A (1937) Fauna marinha de Portugal, 1° aditamento. Memórias e Estud Do Mus Zoológico Da Univ Coimbra 99:1–30
Pallas PS (1771) Reise durch verschiedene Provinzen des Russischen Reichs. Theil 1. Physicalische Reise durch verschiedene Provinzen des Russischen Reichs im 1768- und 1769 sten Jahren. Kayserliche Akademie der Wissenschaften, Saint Petersburg
Pallas PS (1766) Elenchus zoophytorum sistens generum adumbrationes generaliores et specierum cognitarum succintas descriptiones, cum selectis auctorum synonymis. Fransiscum Varrentrapp, Hagae [Den Haag]
Ramil F, Ansín Agís J, Fernández Pulpeiro E (1998) Soft-bottom hydroids (Chidaria: Hydrozoa) collected in the Ría de Vigo (NW Spain). Zool Verh Leiden 323:181–208
Ramil F, Parapar J, Vervoort W (1992) The genus Sertularella Gray, 1848 (Cnidaria: Hydroida) along the coasts of Galicia (Spain). Zool Meded 66:493–524
Ramil F, Vervoort W (1992) Report on the Hydroida collected by the “BALGIM” expedition in and around the Strait of Gibraltar. Zool Verh 277:1–262
Rees WJ, White E (1966) New records and fauna list of hydroids from the Azores. Ann Mag Nat Hist 9:271–284. https://doi.org/10.1080/00222936608656051
Schuchert P (2010) The European athecate hydroids and their medusae (Hydrozoa, Cnidaria): Capitata part 2. Rev Suisse Zool 117:337–555. https://doi.org/10.5962/bhl.part.117793
Schuchert P (2022) World Hydrozoa Database. In: Flanders Mar. Inst. http://www.marinespecies.org/aphia.php?p=taxdetails&id=1337. Accessed 6 Jan 2022
Schuchert P (2012) North-West European athecate hydroids and their medusae. In: Crothers JH, Hayward PJ (eds) Synopses of the British Fauna (New Series), no. 59. Field Studies Council, Linnean Society of London, Estuarine and Coastal Sciences Association, Dorchester, p viii+364
Schuchert P (2004) Revision of the European athecate hydroids and their medusae (Hydrozoa, Cnidaria): Families Oceanidae and Pachycordylidae. Rev Suisse Zool 111:315–369. https://doi.org/10.5962/bhl.part.80242
Servia MJ, Vieira-Lanero R, Cobo F, et al (2006) Notas sobre la presencia de Cordilophora caspia (Pallas, 1771), Dugesia tigrina (Girard, 1850) y Elodea canadensis (Michaux, 1803), en los ríos gallegos. In: EEI 2006–2° Congreso Nacional sobre Especies Exóticas Invasoras (“2nd National Conference on Invasive Alien Species”). GEIB - Grupo Especialista en Invasiones Biológicas, León, p 84
Stechow E (1923) Zur Kenntnis der Hydroidenfauna des Mittelmeeres, Amerikas und anderer Gebiete. II. Teil. Zool Jahrbucher, Abteilung Fur Syst Geogr Und Biol Der Tiere 47:29–270
Svoboda A, Cornelius PFS (1991) The European and Mediterranean species of Aglaophenia (Cnidaria: Hydrozoa). Zool Verh 274:1–72
Teissier G (1922) Observation des médusoides libres et des planules de Sertularia operculata L. Bull La Société Zool Fr 47:357–361
Teissier G (1965) Cnidaires - Cténaires. In: Inventaire de la faune marine de Roscoff - Éditions de la Station Biologique de Roscoff, 2nd edn. Robin & Mareuge, Paris
The Linnean Society (2023) Linnaean Collections. https://www.linnean.org/research-collections/linnaean-collections. Accessed 7 Jan 2023
TriploV (2022) Augusto Pereira Nobre. https://www.triplov.com/biblos/nobre.htm. Accessed 19 May 2022
Universidade do Porto (2022) Augusto Nobre: Publicações, Memória U.Porto. https://sigarra.up.pt/up/pt/web_base.gera_pagina?p_pagina=antigos estudantes ilustres - augusto nobre: publicações. Accessed 19 May 2022
Vervoort W (1993) Report on hydroids (Hydrozoa, Cnidaria) in the collection of the Zoological Museum, University of Tel-Aviv, Israel. Zool Meded 67:537–565
Weber M (1986) Fishing method and seasonal occurrence of glasseels (Anguilla anguilla L) in the Rio Minho, West coast of the Iberian Peninsula. Vie Milieu / Life Environ Obs Océanologique - Lab Arago 36:243–250
WoRMS Editorial Board (2023) World Register of Marine Species. http://www.marinespecies.org/. Accessed 5 Jan 2023
Acknowledgements
The authors acknowledge support provided by Aquamuseu do Rio Minho, municipality of Vila Nova de Cerveira, Portugal. Special thanks to the Research Team of Estuarine Ecology and Biological Invasions (EEBI) of the Interdisciplinary Centre of Marine and Environmental Research (CIIMAR/U.Porto) for collecting samples with the Van Veen grab sampler; to Mafalda Fernandes and Eduardo Martins for sampling via beam trawl fishing; to fisher José Maria via the glass eel fishing bycatch method; and to Harold Cantallo (CIIMAR/U.Porto) for helping collect aquatic plants in the Minho River.
Funding
Open access funding provided by FCT|FCCN (b-on). This study was partially funded by national funds through the Foundation for Science and Technology (FCT, Portugal) within the scope of the project PTDC/BIA-BMA/6363/2020.
Author information
Authors and Affiliations
Contributions
Conceptualization: Dimítri de Araújo Costa; Methodology: Dimítri de Araújo Costa, Carlos Antunes, Nuno Gomes; Formal analysis and investigation: Dimítri de Araújo Costa, Carlos Antunes, Sónia Rocha; Writing—original draft preparation: Dimítri de Araújo Costa; Writing—review and editing: Carlos Antunes, Sónia Rocha, Nuno Gomes; Resources: Carlos Antunes; Supervision: Carlos Antunes.
Corresponding author
Ethics declarations
Competing Interests
The authors disclose that they have no known competing financial interests or personal relationships that could have appeared to influence the study reported in this manuscript.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
de Araújo Costa, D., Antunes, C., Rocha, S. et al. Hydrozoans (Cnidaria) from the estuarine zone of the International Minho River, NW Iberian Peninsula, Atlantic Europe. J Coast Conserv 27, 32 (2023). https://doi.org/10.1007/s11852-023-00965-9
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11852-023-00965-9