Separated by time and depth freshwater Lepidogalaxias is traditionally considered the only member of the family. Now it has a previously overlooked deep sea sister in Malacosteus after testing over 400 competing candidates in the large reptile tree (LRT, 2173 taxa).
Figure 1. Extant Lepidogalaxias to scale with extant Malacosteus, a deep sea relative.
Both have a carnivorous diet, a slender, cylindrical body, needle-like teeth, transparent fin membranes, reddish fixed eyes, and a mobile neck distinct from all other fish (except each other).
Individually the salamanderfish lives in semi-permanent, shallow, acidic pools in Western Australia. It burrows into sand to avoid desiccation.
YouTube video: less than 2 minutes of Lepidogalaxias.
Individually the dragonfish is found worldwide in temperate and tropical seas below 500 meters. It produces red bioluminescence, a color its prey cannot see. Unossified cervical vertetbrae enable great movement. Prey includes zooplankton, which, at depth is 1000x more plentiful than larger items.
YouTube video on Malacosteus.
Malacosteus niger (Ayres 1848, 1849; Kenaley 2007; 25cm) is the extant stoplight loosejaw, a type of deep sea dragonfish, here related most closely to the salamander fish, Lepidogalaxias (above).
Lepidogalaxias salamandroides (Mees 1961; McDowall RM and Pusey BJ 1983; 7cm in length) is the extant salamanderfish, the only fish with a neck capable of turning the head nearly at right angles to the torso. Like lungfish, the freshwater salamanderfish is capable of surviving dry seasons by burrowing into the sand. Note the large eyes and short rostrum. Needle-like teeth line the jaws. Derived from a basal ray fin fish Prohalecites, molecular studies consistently recover Lepidogalaxias close to the base of the Telostei where it also nests in the LRT.
This appears to be a novel hypothesis of interrelationships. If not, please send a citation so I can promote it here.
References Ayres WO 1848. pp. 64–73. In: Proceedings of the Boston Society of Natural History, Vol. 3. Proceedings of the Boston Society of Natural History, Boston. Ayres WO 1849. Description of a new genus of fishes, Malacosteus. Boston Journal of Natural History 6:53–64. Kenaley CP 2007. Revision of the Stoplight Loosejaw Genus Malacosteus (Teleostei: Stomiidae: Malacosteinae), with Description of a New Species from the Temperate Southern Hemisphere and Indian Ocean. Copeia. 2007 (4): 886–900. doi:10.1643/0045-8511(2007)7[886: McDowall RM and Pusey BJ 1983.Lepidogalaxias salamandroides Mees — A redescription, with natural history notes. Rec. West. Aust. Mus. 11(1):11–23. Mees GF 1961. Description of a new fish of the family Galaxiidae from Western Australia. J. Roy. Soc. West. Aust. 44: 33-38.
Co-author, Andrea Cau wrote on Theropoda.blogspot.com: “An element of the fossil that has certainly attracted attention and aroused legitimate online curiosity since its immediate publication is the “crest” that appears on the animal’s snout. Upon immediate, “literal” viewing, Daurlong appears to have a triangular nasal crest running along the dorsal surface of the snout. However, detailed analysis of the fossil and comparison with other specimens disprove this interpretation.”
Andrea Cau wrote this caption to the diagram reproduced below: “Above (Fig 1 below here), Daurlong holotype skull. Below, photo (A) and schematic drawing (B) of the Zhenyuanlong skull. In B, the nasal and part of a lacrimal are indicated in red, the dislocations of which mirror the “crest” of Daurlong.”
Figure 1. The red area is not the nasal and lacrimal, from Andrea Cau. Rather the red area is the dorsal right maxilla and lacrimal. Above, the nasals are in pink/light magenta. Colors are helpful. Measuring the two nasals documents their unequal lengths, equalized by adding the two broken bone chips indicated with arrows.
Andrea Cau wrote on Theropoda.blogspot.com: “Everything leads us to consider the “crest” as an artifact of fossilization, to deny it a biological origin.”
So the crest is not a bone. Is it a stain? Is it paint?
“During the study of the fossil, we authors too have carefully evaluated the hypothesis that the animal had a nasal crest, but in the light of various elements, both geological and biological, we have rejected this interpretation.”
I appreciate that Cau is aware of the controversy and is willing to defend his hypothesis.
Figure 2. Daurlong skull in situ and reconstructed using DGS methods.
Cau continues: “1. The dorsal part of Daurlong’s skull has undergone an erosion of the external surface: this is evident from the analysis with UV rays, which shows a low fluorescence of the dorsal part of the skull compared to the ventral one. The degree of fluorescence of the bones is related to the preservation of the original external surface (more fluorescence = better preservation). The crest is devoid of fluorescence, resulting in deteriorated and eroded bone on the surface. This is also confirmed by sampling of bone material and by electron microscopy analysis.”
The difference is also apparent in white light. Is the crest an artifact of preparation? An unwanted artistic addition? Cau notes only the right ‘crest’ is devoid of fluorescence. If so, why does the left lacrimal rise to the same height? See DGS tracing in figure 2.
“2. The anterior part of the nasal region, at the level of the articulation with the premaxilla, is damaged and missing. It can’t be deduced from the photos, but there is a bony “step” between the “ridge” and the tip of the muzzle, that is, a piece of the lateral part of the premaxilla is missing, the one that articulates with the nasal. This indicates that the original joint between the two bones is missing.”
Again, was novel matter added to this fossil to improve its appeal and price? That’s what we want to know.
Otherwise, tetrapod nasals usually appear in pairs. One or both can leave impressions in the matrix. Are these data dismissed by Cau? Not sure why the pmx-na suture is relevant to the present discussion (But wait! See below).
“3. Another dromaeosaurid from the Jehol Biota, Zhenyuanlong, shows a marked dislocation of the nasals and lacrimals with respect to the rest of the skull: [see Fig 1] this dislocation, evident from the way the specimen is fossilized, produces an “extreme” version of the crest of Daurlong.
As noted above, Cau misidentified the nasal and dorsal arch of the maxilla in Zhenyuanlong (Fig 1). So his example is less convincing than it should be. This is why DGS colors are so important. If you have a leftover bone or two after coloring the easy bones, then get back in there. Those splints and chips need to be colored, too.
“3. Continued… That is, the nasal of the Daurlong specimen is clearly at an early stage in the dislocation process which is more advanced in the Zhenyuanlong specimen.
Now I’m confused. Is the crest “at an early stage in the dislocation” or is it an added artifact that is “devoid of fluorescence.” Cau indicates both are true (perhaps this is a Google translation error).
“3. Continued. The most parsimonious explanation that we get from these elements is that the “ridge” of Daurlong is only the roof of the nasal (and part of the right lacrimal) which partially dislocated dorsally during the decomposition of the carcass. Originally, the roof of the nasal was flat, without ridges.”
Just to back up a bit… We were discussing the right lacrimal ‘crest’. Now Cau turns the argument to the anterior nasal ‘ridge’. Cau reports, “the roof of the nasal was flat.” That is correct. Everyone agrees. That not what we’re wondering about. We are interested in the cresting lacrimals (Fig 2). That vague published outline is intriguing and frustrating. More detailed tracings – and reconstructions based precisely on those tracings, please. No freehand impressions. Stick to the data.
Lacrimal crests are not scored in the LRT. These two taxa (Figs 1, 2) are not dromaeosaurids when the taxon inclusion set is expanded, as it is in the LRT. Toe two is not a killer claw. The pubis was not retroverted.
References Wan X, Cau A, Guo B, Ma F, Qing G and Liu Y 2022. Intestinal preservation in a birdlike dinosaur supports conservatism in digestive canal evolution among theropods. Nature scientific reports. 12:19965 https://doi.org/10.1038/s41598-022-24602-x
Andrea Cau commented, “Your alternative interpretation of Zhenyuanlong skull bones is not valid, because it is contradicted by first hand observation by the authors of its description, and is also challenged by the taphonomic process producing these fossils.”
That could be true. Or not. We’ve seen workers make mistakes before.
“The elements in red are part of the the left nasal and of the right lacrimal, as depicted by the authors who studied the fossil, and is clearly evident from the published photos of the specimen.”
Repeating an argument does not mean you have two arguments. I was hoping Cau would take up the challenge by colorizing all the bones, leaving none omitted.
“I see no reason for trusting your bizarre interpretation of the bones, based just on subjective photoshopping, when the original interpretation by those who studied the fossil is way simpler and more coherent with taphonomy.”
Repeating an argument does not mean you have three arguments. Note, Cau is bringing out demeaning adjectives now. This is unprofessional. Trust is never a part of science. Testing is.
“Both Daurlong and Zhenyuanlong skulls were compressed along the mediolateral direction, and as a consequence the skull roof bones suffered the main dislocation forces: the nasals and lacrimal roofs were dislocated and rotated relative to their original positions, whereas the maxillae suffered only a minimal translation because its main plane was already paralleling that of deposition. You should take carefully into account the taphonomic setting when interpreting a crushed skull.”
Geological and taphonmic backstory is interesting, but we’re looking for all the bones to be identifed. Just to be sure. Not just the red bones.
“The “lacrimal crest” you claimed in Daurlong does not exist: it is just part of the nasal roof, which was dislocated by the taphonomic processes and re-oriented along the main depositional plane. This is a common pattern when a 3-dimensional oreinirostral skull is crushed and compressed by a compression regime oriented along the transverse axis of the skull.”
Quoting the paper:“The skull is almost perfectly articulated, except for the missing nasal ramus of the left premaxilla and the partially displaced dorsal parts of the right nasal and lacrimal.”
“Furthermore, the external surface (periosteal) in most bones of the dorsal half of the skull is eroded, exposing part of the inner bone texture: this is the cause of the different color or the bones, and the reason why some bone boundaries are not more visible: most of the bone shapes in your drawing are just unsupported by direct observation of the fossil under different light conditions.”
Can we just identify ALL the bones, comparing left and right dimensions. Can we build a reconstruction directly from the tracings? Freehand drawings are not a precise with regard to data. What about the left lacrimal of this almost perfectly articulated specimen? Can we identify that?
“In conclusion, there are no “lacrimal crests” in Daurlong, because those elements are just the dorsal roofs of the nasal and lacrimal which were dislocated and exposed along the depositional plane: you misinterpreted a single photo and ignored the taphonomic setting.
Repeating an argument does not mean you have four arguments.
“You should take more attention to taphonomy instead of producing such arbitrary photoshop drawings lacking any empirical support.”
And the demeaning insults make another appearance. Everyone, and I mean everyone uses Photoshop now.
Ironically Cau is trusting a drawing and and ignoring or avoiding the data in the photo.
Earlier the large reptile tree (LRT, 2173 taxa) nested the strange, long-nose Early Devonian ‘placoderm’, Brindabellaspis (Young 1980, Fig 1) with Tiktaalik, a superficially similar, flat-head lobe-fin from the Late Devonian. Back then, in my freshman naiveté, I colored the skull bones following a tetrapod pattern. I even added a hypothetical mandible. That seemed appropriate at the the time, especially given the Middle Devonian tetrapod tracks, but that was a mistake.
Like all fish, some skull bones split while others fuse in patterns that can recall tetrapod homologies. Other times they go their own way. It took time to understand the code, the pattern, the exceptions. After 60 days of housekeeping 414 fish taxa the new tree topology is once again fully resolved.
Recent housekeeping has rescored many taxa, including Brindabellaspis. A new restoration (Fig 1) fills in the missing parts in a basal, jawless placoderm pattern. Only fossil data was scored. DGS colors seem to identify bones faster and easier than abbreviated labels and arrows do when it comes to comparative anatomy and recognizing synapomorphies.
Figure 1. Early Devonian Brindabellaspis now nests with the coeval, but much smaller placoderm, Diadongichthys, in the LRT. Not the similarity in the patterns of lateral lines. Those give a clue as to what bones are present. Brindabellaspis data from Young 1980. Colors added here. Gray ellipses are presumed locations for oral and olfactory openings. The skull is an atrium of gills with openings out the back.
In the end the best match for Brindabellaspis was with the much smaller and much simpler, but coeval Diandongpetalichthys (Fig 1), a taxon that gives us a peek beneath the skull to view the oral and branchial cavity. This skull is essentially an atrium without a floor prior to the invention of jaws. Note the primitive placement of the orbits, visible both dorsally and ventrally, right on the edge, as in finless precursors, like Drepanaspis and Poraspis. You’ll note the gradual lengthening of the rostrum takes place in oly this clade. The other placoderms don’t do this.
Brindabellaspis stensioi (Young 1980; King B, Young GC and Long JA 2018; Early Devonian) was originally considered a strange sort of long-rostrum placoderm. Here it nests with Diandongichthys, another flat-head placoderm. The wrap-around nasal and jugal are restored here.
Diandongpetalichthys liaojiaoshanensis (Zhu et al. 1991; Early Devonian) is a petalichthid placoderm close to Lunaspis. The postorbital is restricted to the lower orbit. Jaws are lacking.
Rarely do reference lists include authors whose names begin only with Y and Z. Today is one of those rare days.
References Young GC 1980. A new Early Devonian placoderm from New South Wales, Australia, with a discussion of placoderm phylogeny. Palaeontogr. Abt. A Palaeozool-Stratigr. 167: 10–76. Zhu M 1991. New information on Diandongpetalichthys (Placodermi: Petalichthyida). Early vertebrates and related problems of evolutionary biology. Beijing: Science Press. 179–192. Zhu et al. 2012. An antiarch placoderm shows that pelvic girdles arose at the root of jawed vertebrates. Biology Letters Palaeontology 8:453–456.
In birds, bats, most pterosaurs (sans bottom-decker tapejarids) and flying fish (Figs 1, 2) the wings and homologous pectoral fins are raised to the dorsal half of the torso (Figs 2, 3) by a taller coracoid or its equivalent (e.g. the clavicle in bats).
Figure 1. Flying fish (Exocoetus volitans) line drawing. Note the elevated insertion of the pectoral fins. This expands the size of the pectoral muscles. Despite this, flying fish do not flap when they fly.
That’s not the case with the several so-called flying fish of the Triassic like Thoracopterus (Fig 3) and Potanichthys (Fig 3). Instead, the oversized pectoral fins were anchored low on the torso (Fig 1), more like the extant flying gurnard, Dactylopterus (Fig 3).
Figure 2. Exocoetus skull and pectoral girdle. Note the high placement of the pectoral fins on this fish capable of gliding, but not flapping in the air.
Recent housekeeping in the large reptile tree (LRT, 2173 taxa) now nests Dactylopterus with Thoracopterus and Potanicthys. Everyone agrees that the latter two were not related to extant flying fish, but apparently no published materials have linked these three taxa together phylogenetically.
Figure 3. Dactylopterus, the flying gurnard, along with related taxa, including Thoracopterus and Potanichthys. Parasemionotus lacks enlarged pectoral fins, but was part of the same radiation.
Dactylopterus never breaks the surface to fly or glide. Rather it is a bottom dweller (see video below), moving like vaguely similar bottom-dwelling skates and rays. Look closely and you’ll see Dactylopterus elevates itself off the sea floor by standing, walking and running on is pelvic fins. Yes, it’s another biped with wings, like birds (and kin), Microraptor, Zhenyuanlong, basal pterosaurs (and kin).
YouTube video of Dactylopterus walking on its pelvic fins (making it an underwater biped!) and spreading its pectoral fins as a bottom-dwller, not a flying fish. Colorful patterns like this are not found on the wings of flying fish, suggesting a decorative or flash signal use for these oversize pectoral fins.
And if you’re curious about the origin of flying fish… in the LRT flying fish (Exocoetus) evolved from jacks (Seriola, Fig 4).
Figure 4. Seriola and Exocoetus to scale.
Xu and Zhao 2015 discussed the origin of the Thoracopteridae (Fig 5). These authors also found Peltopleurus in the lineage of Triassic fish with over-sized pectoral fins.
Figure 5. From Xu and Zhao 2015, rotated and colors added here. Peltopleurus nests here in the ancestry of Triassic fish with over-sized wings. Dactylopterus is missing.
Otherwise, these appear to be novel hypotheses of interrelationships. If not, please provide a citation so I can promote it here.
References Xu G-H and Zhao L-J 2015. From Potanichthys to Wushaichthys: resolvingthe evolutionary origin and reproductive strategy of the Thoracopteridae: a reply to Tintori (2015). Biol. Lett. 11: 20150604. http://dx.doi.org/10.1098/rsbl.2015.0604
Qiao et al 2022 report, “Omphalosauridae, comprising several species in the genus Omphalosaurus, is an enigmatic group of durophagous marine reptiles known hitherto from Early–Middle Triassic pelagic sediments of Western North America, Svalbard, and the Bavarian Alps”.
The authors make the following citations: “Omphalosauridae Merriam, 1906 (= Nasorostra Jiang et al., 2016)”. The authors nest this clade within the Ichthyosauriformes and within the Ichthryosauriformes within the clade Diapsida.
Qiao et al define Omphalosauridae (Merriam 1906) as: “The last common ancestor of Omphalosaurus nevadanus, Cartorhynchus lenticarpus and Sclerocormus parviceps and all of its descendants.”
In the LRT (Fig 3) that last common ancestor is Claudiosaurus, a marine younginiform. “All descendants” in the LRT include sauropterygians, ichthyosaurs, mesosaurs, thalattosaurs and the omphalosaur clade, which is not the authors’ intent. Basically this new clade definition is Enaliosauria + Claudiosaurus and the Omphalosaurus clade slips in between them. That’s OK.
Taxon exclusion mars this report. At least two clades of unrelated taxa have a diapsid skull in the large reptile tree (LRT, 2173 taxa). Qiao et al did not know this. Ichthyosaurs nest with Sauropterygia within the Enaliosauria. Qiao et al are not aware of this. That’s why adding taxa is so important. Cherry-picking taxa leads to false positives.
Figure 1. Omphalosaurus nevadanus, MBG 1500, the Austria specimen, skull portion in situ, mandible parts, skull parts and reconstruction with matching colors based on Sinosaurosphargis, the only other taxon with such a large set of splenials.
In the LRT Omphalosaurusnests in a clade basal to the Enaliosauria (Fig 3). Other clade members include Atopodentatus, Sinosaurosphargis, Largocephalosaurus. The proximal outgroup taxon is Claudiosaurus. Only two of these were mentioned and tested in the Qiao et al analysis: Sinosaurosphargis and Largocephalosaurus.
Figure 2. Cladogram from Qiao et al 2022 showing the taxon exclusion and cherry picking of taxa. These authors have no idea how these taxa are interrelated due to their omission of many taxa.
Qiao et al report, “Here we report the large basal ichthyosauriform Sclerocormus from the Early Triassic of China that clearly demonstrates an omphalosaurid affinity, allowing for the synonymy of the recently erected Nasorostra with Omphalosauridae.”
That hypothesis of interrelationships is not supported by the LRT (Fig 3).
Figure 3. Subset of the LRT focusing on the Enaliosauria with emphasis on Omphalosaurus and Sclerocormus. Adding taxa moves these two away from ichthyosaurs.
The authors mistakenly consider Sclerocormus and Cartorhynchusto be members of the Ichthyosauriformes. Adding taxa moves both to the base of the Sauropterygia close to Qianxisaurus, yet another taxon missing from the authors’ analysis.
References Qiao Y et al (7 co-authors) 2022. A globally distributed durophagous marine reptile clade supports the rapid recovery of pelagic ecosystems after the Permo-Triassic mass extinction. Nature communications biology https://doi.org/10.1038/s42003-022-04162-6
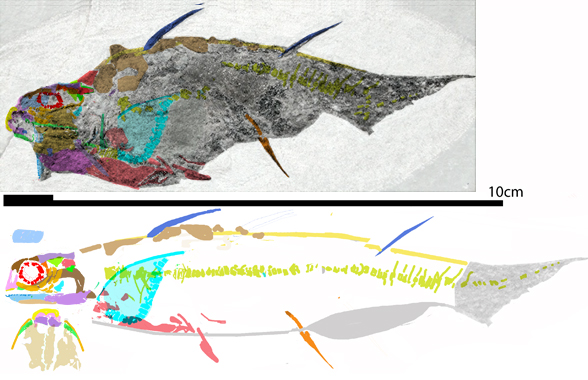
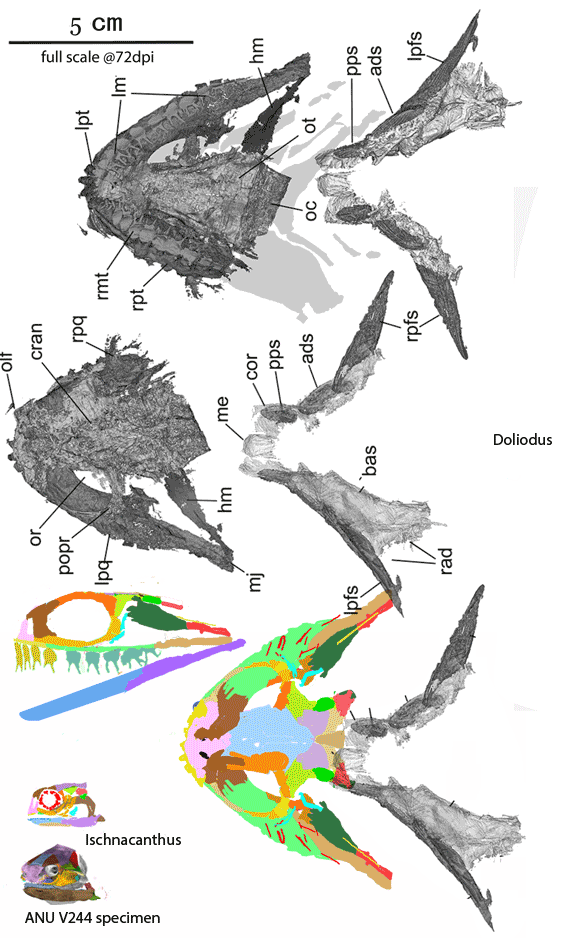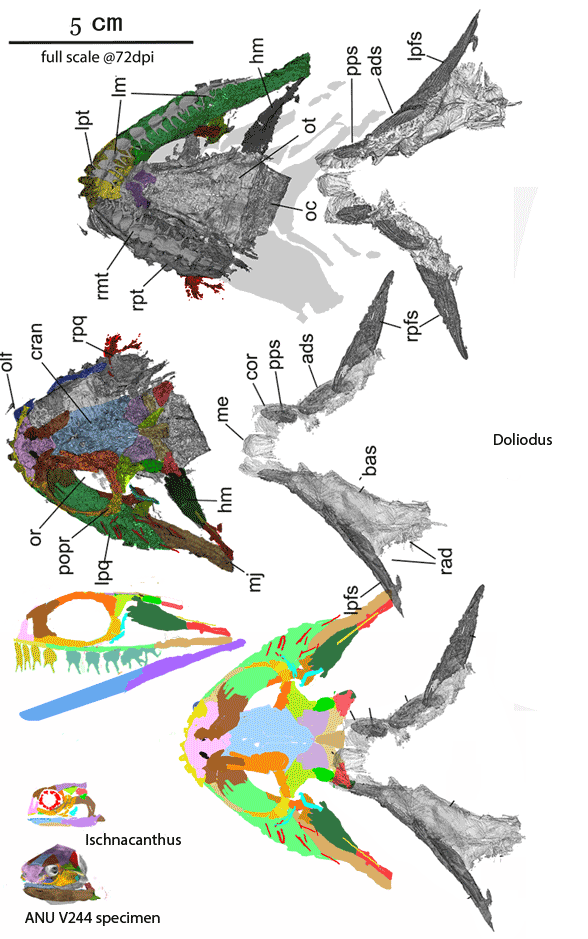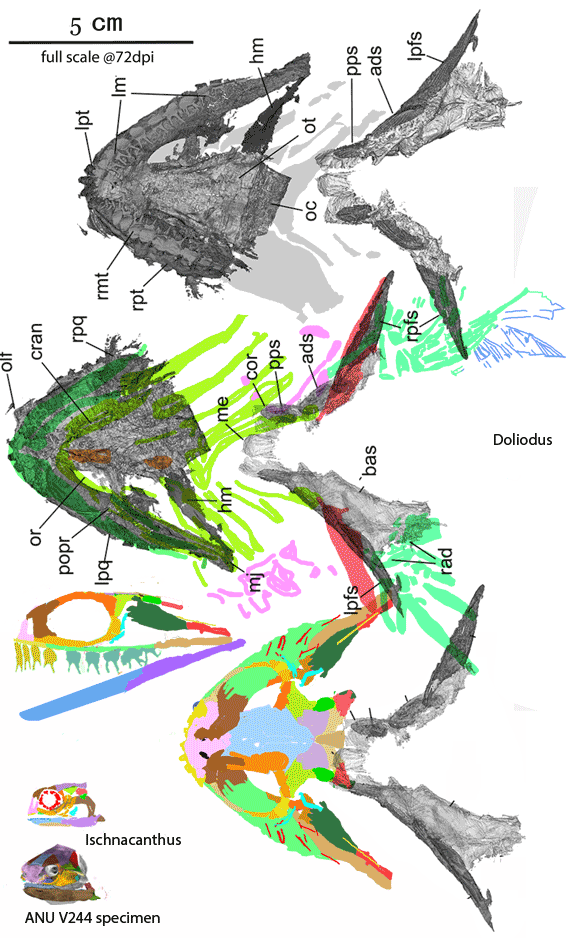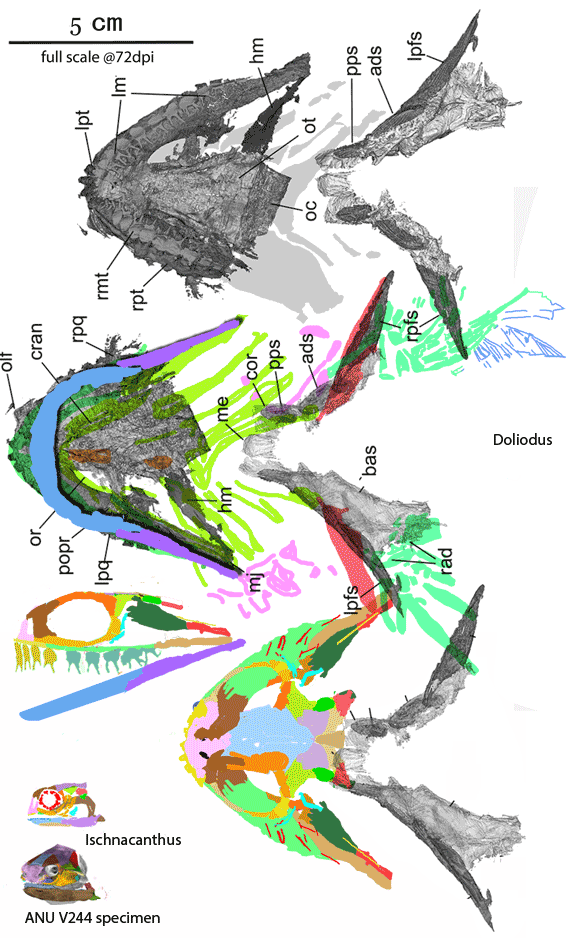
Recent housekeeping in the fish subset of the large reptile tree (LRT, 2173 taxa, Fig 5) sheds new light on this newly revised DGS tracing of the crushed skull of the Early Devonian spiny shark, Ischnacanthus (Figs 1, 2). The palate is visible here. That’s rare and attests to the relatively wide skull, wider than in most spiny sharks. That enabled comparison with the palate of the Unnnamed ‘arthrodire placoderm’ with the museum number ANU V244 (Fig 1). Turns out the two are quite similar.
Figure 1. Tiny taxa at the transition between placoderms and spiny sharks: Early Devonian ANU V244 (still unnamed) and Early Devonian Ischnacanthus. Other than the skull, the ANU V244 specimen preserves a placoderm-like ventral plate, but nothing else of the post-crania is known, including the fins. Colors added here. ANU V244 photos from Hu, Lu and Young 2017.
Tiny Early Silurian Shenacanthus (Fig 3) is ancestral to both the Early Devonian ANU V244 specimen and coeval Ischnacanthus.
The origin of teeth In these DGS tracings (Figs 1–3) DGS colors are applied with tetrapod homologs. The green patches with bumpy ventral surfaces are maxillae with maxillary teeth. That’s how teeth first appear in vertebraes. In these taxa maxillary teeth are not yet on the margin, but hidden by postorbital cheeks. The yellow patch below the nasal is the premaxilla and its carpet of tiny bumpy teeth. The large tan arches are lacrimals, anchoring the maxillae. Lacrimals are associated with respiration now and so are restricted to the narial area, but in fish respiration occurs at the gills. That’s why lacrimals first appeared below the braincase and in back of the skull. The blue dentaries lie loosely on the mandible. The lavender patches in the palate are vomers, also provided with tiny tooth carpets.
Figure 2. Ischnacanthus in situ and reconstructed, both about 1.3x life size.
A related tiny taxon, Shenacanthus vermiformis (Zhu et al 2022, Early Silurian, est 3cm long, Fig 3) is the earliest known of the jawed placoderms and the basalmost member of the spiny sharks. Claspers are present. So are lacrimals and maxillae.
Unnnamed specimen (ANU V244, Young, Lelièvre and Goujet 2001; Hu, Lu and Young 2017; Early Devonian (Fig 1) is a tiny relative to Coccosteus and even tinier Shenacanthus).
Ischnacanthus gracilis (Egerton 1861, orginally Diplacanthus gracilis, Burrow et al 2018; Miles 1973; Early Devonian, 410 mya; up to 2m in length, Figs 1, 2) is an early acanthodian with placoderm-like maxillary pads and a wide skull based on the wide sphenoid (braincase floor), as in the ANU V244 specimen.
Figure 3. Early Silurian Shenacanthus is no more than 3cm in length, basal to all bony fish including the ANU V244 specimen, Ischnacanthus, tetrapods and humans.
The Late Silurian placoderm, Entelognathus(Zhu et al. 2013), now nests at the base of the Chondrichyes (sharks, rays, ratfish) in the LRT.
The arthrodire placoderm, Coccosteus. is the last common ancestor of sharks and spiny sharks + bony fish in the LRT (Fig 5). This pushes the origin of placoderms into the Ordovician, prior to the appearance of Early Silurian Shenacanthus, a phylogenetically miniaturized taxon (Fig 3).
Figure 4. Doliodus and Ischnacanthus are sisters in the LRT now. Several spines bind together here to form a robust spine at the anterior margin of the broad pectoral fins, convergent with Iniopterygia. The robust teeth have twin cusps, convergent with the basal chondrichthyan, Xenacanthus and all point medially (not ventrally as shown for clarity in the lateral view. The maxillae are longer and the lacrimals are flatter.
Somewhat larger Doliodus also joins Ischnacanthus (Fig 4) at the base of the spiny sharks in the LRT (Fig 5). Several pectoral spines = rays bind together here to form a robust spine at the anterior margin of the broad pectoral fins, convergent with Iniopterygia and catfish. The robust teeth have twin cusps, convergent with the basal chondrichthyan, Xenacanthus, but all teeth lay flat and point medially. The maxillae are longer than in Ischnacanthus and the lacrimals are flatter. As in Climatius and Brachygnathus (Fig 3), several small post-pectoral spines are present along the ventral abdomen.
Figure 5. Subset of the LRT focusing on basal vertebrates as of 12.2022.
This appears to be a novel hypothesis of interrelationships. If not, please provide a citation so I can promote it here.
References Burrow CJ et al. 2018. The Early Devonian ischnacanthiform acanthodian Ischnacanthus gracilis (Egerton, 1861) from the Midland Valley of Scotland. Acta Geologica Polonica 68(3): 335–362. Egerton P de MG 1861. British fossils. (Descriptions of Tristichopterus, Acanthodes, Climatius, Diplacanthus, Cheiracanthus). Memoirs of the Geological Survey of the United Kingdom (British Organic Remains), Decade 10: 51–75. Hu Y, Lu J and Young 2017. New findings in a 400 million-year-old Devonian placoderm shed light on jaw structure and function in basal gnathostomes. Nature Scientific Reports 7: 7813 DOI:10.1038/s41598-017-07674-y Miles R 1973. Articulated acanthodian fishes from the Old Red Sandstone of England, with a review of the structure and evolution of the acanthodian shoulder-girdle. Bulletin of the British Museum (Natural History), 24, 111–213. Young G C, Lelièvre H and Goujet D 2001. Primitive jaw structure in an articulated brachythoracid arthrodire (placoderm fish; Early Devonian) from southeastern Australia. J. Vertebr. Paleontol. 21, 670–678. Zhu Y-A et al (10 co-authors) 2022. The oldest complete jawed vertebrates from the early Silurian of China. Nature 609:954–958. online
wiki/Ischnacanthus wiki/Shenacanthus – not yet posted wiki/ANU v244 not yet posted, not yet named
An alternate view from eleven years ago: Blais S, MacKenzie L and Wilson M 2011. Toothlike scales in Early Devonian eugnathostomes and the ‘outside-in’ hypothesis for the origins of teeth in vertebrates Journal of Vertebrate Paleontology, 31 (6), 1189-1199 DOI: 10.1080/02724634.2011.607992
According to the LRT, teeth arose much earlier, in the Early Silurian, when placoderms gave rise to sharks and spiny sharks.
Ongoing housekeeping to the fish subset of the large reptile tree (LRT, 2173 taxa) moves the extant viperfish (Chaulodus, Fig 1) closer to Middle Triassic to Early Cretaceous Leptolepis and Middle Triassic Prohalecites.
Figure 1. The viperfish, Chauliodus, measures 60cm. Here shown to scale with Mesozoic relatives Leptolepis and Prohalecites.
Chauliodus sloani (Forster in Bloch and Schneider 1801, up to 60cm in length) is the extant viperfish. A tiny glowing lure from the anterior dorsal fin lure deep sea fish to the oversized teeth. Scales and maxillary teeth are retained. Apparently the temporal series were not retained. The anterior dorsal fin is new.
Figure 2. Leptolepis skull in situ and slightly reconstructed.
The large eyes and absent circumorbital bones of Chauliodus were inherited from its smaller ancestor, Leptolepis (Fig 2). The long teeth and lure evolved later.
References Arratia G and Tintori A 1999. The caudal skeleton of the Triassic actinopterygian †Prohalecites and its phylogenetic position, p. 121–142. In: Mesozoic Fishes 2—Systematics and Fossil Record. G. Arratia and H.-P. Schultze (eds.). Verlag Dr. F. Pfeil, München. Arratia G 2015. Complexities of early Teleostei and the evolution of particular morphological structures through time. Copeia 103(4):999–1025. Bellotti C 1857. Descizione di alcune nuove specie di pesci fossili di Perledo e di altre localtta lombarde. 419–432. In Sopani A (ed) Studi geologici sulla Lomabardia. Editore Turati, Milano. Forster JR 1801. in Bloch ME and Schneider JG editors, Systema Ichthyologiae Iconibus cx Ilustratum. Post obitum auctoris opus inchoatum absolvit, correxit, interpolavit Jo. Gottlob Schneider, Saxo. Berolini. Sumtibus Auctoris Impressum et Bibliopolio Sanderiano Commissum. i-lx + 1-584. Tintori A 1990. The actinopterygian fish Prohalecites from the Triassic of northern Italy. Palaeontology 33:155–174.
Goswami et al 2022 reported, “While molecular data has largely resolved the higher-level topology of placental mammals and there are detailed morphological phylogenies available for many extinct placental clades, there is continuing debate over the positions of some entirely extinct clades within Placentalia.”
Sadly, like many paleontologists nowadays, the authors drank the Kool-Aid. They believed in deep time genomic results even though their cladogram nested mysticetes with odontocetes and elephants as basal placentals. Their ingroup includes members of the clade Creodonta, which the large reptile tree (LRT, 2173 taxa) nest within Marsupialia. The taxon, Monodelphis, (the marsupial wthout a pouch nesting as the proximal outgroup to the Placentalia in the LRT) is not mentioned in the SuppData nor tested in their cladogram.
A wider gamut of taxa in the LRT tells us which taxa are ingroup members and outgroup members. Cherry-picking taxa (all too common in paleontology) does not work as well.
From their abstract: “Analysis of a three-dimensional skull dataset [Fig 1] for living and extinct placental mammals demonstrates that evolutionary rates peak early and attenuate quickly. This long-term decline in tempo is punctuated by bursts of innovation that decreased in amplitude over the past 66 million years. Social, precocial, aquatic, and herbivorous species evolve fastest, especially whales, elephants, sirenians, and extinct ungulates. Slow rates in rodents and bats indicate dissociation of taxonomic and morphological diversification.”
Since these authors have no idea what the actual topology of Placentalia is, none of their hypotheses have anything to do with reality. The “peak early and attenuate quickly” hypothesis is not supported by the LRT. See below.
The authors tested skull shape only and applied shape coordinates (Fig 1). I was curious if they tested Mesonyx and Hippopotamus, two herbivores with large fangs. They did not. One wonders if they were deleted because they nested with other placentals with large fangs?
In the Goswami et al study, at the base of whales is Elotherium (aka Entelodon, a giant wart hog-like artiodactyl of the Eocene to Miocene. It’s going to be hard to find the transitional taxa linking a taxon that has hooves and reduced digits to mysticetes + odontocetes which do neither in their flippers. To the Goswami team, the editors and referees that doesn’t matter. Elsewhere in their cladogram bats nest with hedge hogs, moles and shrews. None of these are arboreal. Or leap. Or chirp. Or have long limbs. Or primitive skulls.
By contrast in the LRT all sister taxa and clades grade gradually from one to another, modeling actual evolutionary events.
Figure 2. Skeleton of Tenrec alongside restored skeleton model of Pakicetus. Despite these two nearly identical (apart from size) skeletons and skulls, Goswami et al nested Tenrec near basal placentals and Pakicetus about as far as possible away on their cladogram. Just one of many obvious blunders.
What the LRT recovers: Placentalia appeared in the Early Jurassic. Taxa remained small, arboreal and nocturnal until the dinosaur extinction event. Glires (including Jurassic Mutituberculata), primates and leptictids all have Mesozoic roots. Almost immediately after the dinosaurs larger descendants appeared and radiated into diurnal and terrestrial taxa. That means throughout the Jurassic and Cretaceous (2/3 of their time on Earth) placentals did not diversify much, contra the Goswami et al hypothesis.
References Goswami A et al (12 co-authors) 2022. Attenuated evolution of mammals through the Cenozoic. Science 378(6618(:377–383.
Publicity Santana SE and Grossnickle DM 2022. Bursts in skull evolution weakened with time. Science 378(6618(:355–356.
Santana and Grossnickle 2022 reported, “Evolutionary biologists generally agree that placental mammals began as small-sized animals that ate insects, and later evolved to become more varied in size and morphology.
That’s not what Goswami et al recovered. Just the opposite.
“Today, the morphologies of placental mammals are incredibly diverse, ranging from the bumblebee bat to the blue whale. There is considerable debate over how the small, insect-eating ancestors of placental mammals gave rise to such incredibly divergent lineages.
That’s what LRT models, subject to confirmation, refutation and/or modification.
“Among the competing hypotheses are varying opinions on the timing and pace of early placental mammal evolution, especially in the context of major environmental changes such as the Cretaceous-Paleogene (K-Pg) mass extinction event 66 million years ago and the Paleocene-Eocene Thermal Maximum (PETM) 56 million years ago. On page 377 of this issue, Goswami et al. (4) contribute to this discussion by reconstructing the patterns and possible drivers of placental mammal morphological diversification with a quantitative analysis of skull shape spanning over 70 million years of evolution.”
First start with a suitable outgroup recovered by builiding your own LRT. Then include as many taxa as possible until the evolutionary patterns emerge from the convergent exceptions.
Wan et al 2022 report, “Dromaeosaurids were bird-like dinosaurs with a predatory ecology known to forage on fish, mammals and other dinosaurs. We describe Daurlong wangi gen. et sp. nov., a dromaeosaurid from the Lower Cretaceous Jehol Biota of Inner Mongolia, China.”
Like Wan et al,the large reptile tree (LRT, 2173 taxa) also nests Daurlong with Zhenyuanlong but in the lineage of Tyrannosaurus, with many traits convergent with dromaeosaurids, according to the LRT. Taxon exclusion mars the Wan et al theropod study.
Figure 1. Zhenyuanlong (above) compared to scale with Daurlong (below). The cervicals are too small and too long in the freehand graphic by Wan et al. There is no indication of a killer claw in the pes of Daurlong. The pubis should be directed ventrally, not posteroventrally.
Wan et al 2022 continue, “Exceptional preservation in this specimen includes a large bluish layer in the abdomen which represents one of the few occurrences of intestinal remnants among non-avian dinosaurs. Phylogenetically, Daurlong nests among a lineage of short-armed Jehol Biota species closer to eudromaeosaurs than microraptorines.”
That may be due to taxon exclusion. Huaxiagnathus is the outgroup taxon in the LRT, but not included in the Wan et al study. Many other taxa are needed to separate tyrannosaur ancestors from dromaeosaurids.
Figure 2. Daurlong skull in situ and reconstructed using DGS methods. DGS methods help recover more data. Others are using DGS in µCT studies.
Wan et al 2022 continue, “The topographic correspondence between the exceptionally preserved intestine in the more stem-ward Scipionyx and the remnants in the more birdlike Daurlong provides a phylogenetic framework for inferring intestine tract extent in other theropods lacking fossilized visceral tissues. Gastrointestinal organization results conservative among faunivorous dinosaurs, with the evolution of a bird-like alimentary canal restricted to avialan theropods.”
The intestinal preservation is interesting, but the phylogenetic context needs more taxa. Figure 2 indicates relatively little attention to detail was undertaken to identify skull elements by Wan et al and the authors assumed a dromaeosaurid hypothesis by focusing on a few traits. We call this “Pulling a Larry Martin“. Minimize taxon omission to avoid this problem.
That’s why the authors gave Daurlong a ‘killer claw’ and retroverted pubis it did not have. That’s why the ignored the twin lacrimal crests and the more robust cervicals.
As Daurlong and Zhenyuanlong demonstrate tyrannosaur ancestors were also bird-like dinosaurs.
Added Dec 2, 2022: about those crests, click here.
References Wan X, Cau A, Guo B, Ma F, Qing G and Liu Y 2022. Intestinal preservation in a birdlike dinosaur supports conservatism in digestive canal evolution among theropods. Nature scientific reports. 12:19965 https://doi.org/10.1038/s41598-022-24602-x
Figure 1. Subset of the LRT focusing on Sauropterygia. Here Hanosaurus nests well inside Sauropterygia.
The Wang et al November 2022 paper, like its May 2022 predecessor, suffered from taxon exclusion. described earlier here..
Figure 2. Frame 1: Cladogram from Wang et al. May 2022. Frame 2: Color overlay indicates clades in the LRT. Blue/Cyan = Ichthyopterygia. Violet/Lavendar = Thalattosauria. Pink = Eosauropterygia. Yellow/green = Placodontia. This Wang et al. cladogram could use more ingroup and outgroup taxa. The ‘ancestor’ is Youngina capensis. Compare to a subset of the LRT in figure 1.
References Riepppel O 1998. THe systematic status of Hanosaurus hupehensis (Reptilia, Sauropterygia) from the Triassic of China. Journal of Vertebrate Paleontology 18(3):545-557. Wang W, Shang Q, Cheng L, Wu X-C and Li C May 2022. Ancestral body plan and adaptive radiation of sauropterygian marine reptiles. bioRxiv preprint doi: https://doi.org/10.1101/2022.04.25.489368 Wang W, Shang Q, Cheng L, Wu X-C and Li C November 2022. Ftable Ancestral Body Plan and Adaptive Radiation of Sauropterygian Marine Reptiles, ISCIENCE (2022), doi: https:// doi.org/10.1016/j.isci.2022.105635. Young CC 1972. A marine lizard fron Nanchung, Hupeh province. Memoirs of the Institute of Vertebrate Paleontology and Paleoanthropology. Academia Sinica 9:17-28.