Daniel Paluh (U Florida) 2022 presented a comprehensive seminar on frogs in the YouTube Royal Tyrrell Museum of Paleontology series. Well worth watching. No taxon exclusion here! This is a master class on frogs.
Paluh describes ‘hyper-ossification’ in certain frog skulls That should be seen in the context of hypo-ossification for frogs in general (Fig 1). Adding bone to a hypo-ossified skull to make it hyper-ossified could be considered a reversal to more highly ossified frog ancestors like Micromelerpeton and Doleserpeton(Fig 1), except that’s not quite the case. Paluh shows how some hyperossified frog skulls are used to plug burrow entrances. These frogs migrate the eyeballs laterally, distinct from typical pond frogs with dorsal orbits for peering out over the surface and distinct from precursor taxa.
Figure 1. Skulls in several views of frog ancestors rom Micromelerpeton, Doleserpeton, Batrachognathus and Rana, documenting skull hypo-ossification during phylogeny.
What is a frog? Essentially a frog is a salamander with a short backbone, long pelvis, long legs, large hands and extra large feet (Fig 2), all of which arise from an increasingly effective hopping behavior. As Paluh shows, dentary teeth disappear 20x by convergence. This likely occurred as the soft, stretchy, sticky, fast-extending frog tongue was used to capture prey and pull it back into the wide jaws. That seems obvious in hindsight, but fascinating to hear and understand for the first time.
Figure 2. Evolution of frogs to scale from Doelserpeton through Gerobatrachus, Triadobatrachus and Rana. Click to enlarge.
Highly recommended YouTube. Daniel J Paluh’s results were recovered after years of work looking at hundreds of frogs.
From Paluh’s website: “I am an NSF PRFB postdoctoral researcher at the University of Florida and Harvard University investigating the molecular and developmental mechanisms of convergent tooth loss in frogs. My research aims to link phylogenetic patterns and underlying developmental mechanisms to better understand the evolutionary processes that generate phenotypic variation in vertebrates. My work integrates micro-computed tomography scanning, comparative methods, and developmental genetics to test macroevolutionary questions. I recently completed my PhD at the University of Florida and Florida Museum of Natural History.”
Once again, taxon exclusion ruins an otherwise perfectly good paper about an important specimen (Figs. 1, 2) that looks like what it is not.
Figure 1. Honghesaurus in situ from Xu et al. 2022 shown < half size. Total length = 47cm or 18.5 inches.
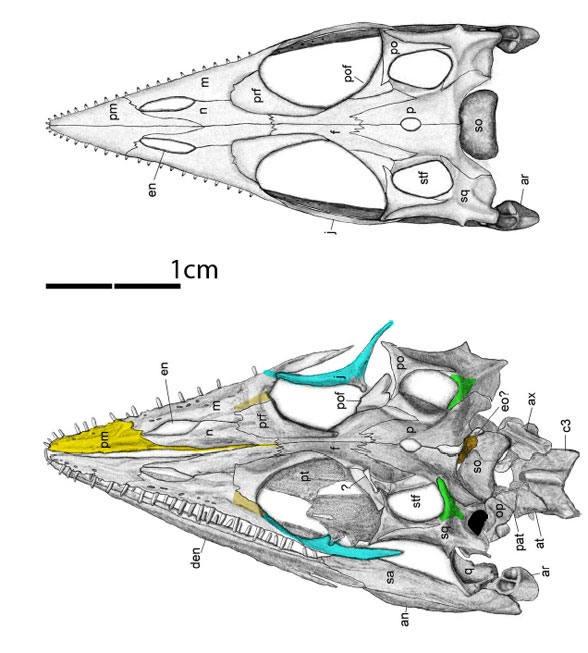
Honghesaurus longicaudalis (Xu et al. 2022, Middle Triassic, IVPP V30380) was considered a pachypleurosaur, but here in the large reptile tree (LRT, 2085 taxa; Fig 3) nests as a transitional taxon betweenAnarosaurus and (mesosaurs + thalattosaurs) + (Wumengosaurus + ichthyosaurs) distinct from the pachypleurosaur lineage.
Figure 2. Skull of Honghesaurus in situ and reconstructed from Xu et al. 2022. The authors missed the longer ascending process of the premaxilla (yellow), the lacrimals (light tan), the anterior jugals (cyan), the supratemporals (green) and the postparietal (dark tan) that fits into the slot between the parietals when not crushed. The mandibles should be wider than the temples in vivo.
Lesson #1 to learn here: Run your analysis before you assume what seems obvious. Honghesaurus looks superficially like a pachypleurosaur. But it doesn’t nest with pachypleurosaurs. It’s the first splinter off the pachypleurosaurs basal to other clade of enaliosaurs. That’s going to happen every so often. This is one of those long-sought transitional taxa. This is what the last common ancestor of mesosaurs, thalattosaurs, Wumengosaurus and ichthyosaurs looks like. It looks like a pachypleurosaur, but is not one.
Figure 3. Subset of the LRT focusing on basal enaliosauria. Xu et al. omitted many of the pertinent taxa, including outgroup taxa, which is why their cladogram does not match this one.
Lesson #2 to learn here: Xu et al. 2022 provide two cladograms (Fig 4). One rooted by placodonts. The other rooted by Youngina. The authors are telling us they have don’t know which one is correct. Don’t make the same mistake. Before you send your manuscript off, add so many taxa to your own LRT that your software will tell you which outgroups are correct and which taxa are pertinent ingroup taxa.
Figure 4. Two cladograms from Xu et al. 2022 each rooted by a different set of outgroup taxa demonstrating the authors have no idea which outgroup is correct. Don’t do this. Find out by creating your own LRT.
Xu et al. is a cautionary tale literally telling fellow workers they are driving blindfolded. This self-confessed problem is also another black mark on Nature and other academic publishers, its editors and referees who found this basic error acceptable. They should have sent it back to the authors, telling them to add taxa to their taxon list. Unfortunately, this is the state of paleontology today. We’ve seen this sort of taxon omission often, as longtime readers know all too well. To academics it’s OK to not know, and to not want to know, even though non-academic scientists were able to figure this out and put it online years ago.
Honghesaurus is a pristine perfect and important specimen, but not for the reasons Xu et al. present. They wrote, “Pachypleurosaurs (Pachypleurosauroidea) are a group of small to medium-sized, lizard-like marine reptiles in the Early to Middle Triassic, including Pachypleurosauridae, Keichousauridae and closely related taxa. The group is generally considered as a sauropterygian radiation, but its phylogenetic interrelationships remain highly debated.”
‘Highly debated’ is hyperbole, meant to make this paper sound more important. Yes, pachypleurosaurs are sauropterygians, but Honghesaurus is not. So-called debates like this are always settled by adding taxa.
“Honghesaurus, although possessing a series of autapomorphies, fills the morphological gap between Qianxisaurus from the Ladinian Xingyi Biota and Wumengosaurus from the Anisian Panxian Biota.”
Likewise, Honghesaurus nests between the pachypleurosaur, Qianxisaurus and the basal ichthyosaur, Wumengosaurus in the LRT (Fig 3). But look how many taxa Xu et al. overlooked in their cladogram (Fig 4) relative to the taxon list in the LRT (Fig 3). Due to taxon exclusion the authors had no idea how important their new enaliosaur was in a phylogenetic context. They could have solved several long-standing problems simply by adding taxa to their cladogram.
Given the phylogenetic node on which Honghesaurus rests, (Fig. 3) this taxon had its genesis in the Early Permian, prior to the appearance of Early Permian mesosaurs.
Congratulations to the authors. Thank you for publishing on Honghesaurus. Please add taxa to your next analysis. Don’t assume a taxon list. Don’t cherry-pick taxa. Don’t borrow a cladogram. Build your own. Give it so many taxa that it will tell you which taxa are outgroups and which are ingroups.
PS A professional note from the lead author arrived the same day: “Dear David, I am fully considering your suggestions. Thank you for your helpful comments and I will add more taxa to reassess the phylogenetic position of Honghesaurus in my next analysis. Please keep in tough [= touch]. Best wishes, Guang-Hui
References Xu G-H, Ren Y, Zhao L-J, Lioa J-Land Feng D-H 2022. A lon-tailed marine reptile from China provides new insights into the Middle Triassic pachypleurosaur radiation. Nature Scientific Reports 12:7396. https://doi.org/10.1038/s41598-022-11309-2
From the Wang et al. abstract: “Here, the pectoral girdles of the Early Cretaceous birds Sapeornis and Piscivorenantiornis are reconstructed for the first time based on computed tomography and three-dimensional visualization, revealing key morphological details that are important for our understanding of early-flight evolution. Our study reveals additional lineage-specific variations in pectoral girdle anatomy, as well as significant modification of the pectoral girdle along the line to crown birds.”
Figure 1. Figure from Wang et al. 2022. Animation and arrows added here. Note the elongation of the locked down coracoids, raising the shoulders above the midline. Microraptor is convergent with birds.
Figure 2. Piscivorenantiornis from 1.18.22. Note the locked-down elongate coracoids, indicating a flapping capability.
Wang et al focused on the details and overlooked the basics, the overall bauplan. In basal theropods, as in basal tetrapods, the short, round coracoids slid along the rims of the small sternum (Fig 1 top). In flapping taxa, like Microraptor and Columba (Fig 1) the coracoid stops sliding, is locked down and elongates raising the shoulders above the midline.
Figure 3. Sapeornis specimen STM 15-15 reconstructed from DGS tracing, figure 1 compared to a more robust specimen with larger feet but an identical humerus. Here the clavicles are shorter with rounder verntral margins, indicating a reduced flapping ability, despite the large wings.
This bauplan is repeated in pre-pterosaurs, like Cosesaurus using the same elements by convergence.
Figure 4. Cosesaurus flapping. Note the locked down elongate coracoids.
This flapping bauplan is also repeated in bats using a locked down and elongate clavicle instead of a coracoid, missing mammals.
Figure 5. Ptilcercus (above) and Icaronycteris (below). Note the ellongate clavicle in the bat, raising the shoulders to the dorsum.
Wang et al. make no mention of nonvolant flapping. As everyone knows, most tetrapods, including gliding taxa, advance the forelimbs left-right-left-right. Flappers don’t. They flap symmetrically. Thus bipedalism is a precursor configuration required for flapping.
Are bats bipedal? Yes, they are inverted bipeds, hanging by their toes.
References Wang et al. 2022. Digital restoration of the pectoral girdles of two Early Cretaceous birds and implications for early-flight evolution. eLife 2022;0:e76086. DOI: https:// doi. org/ 10. 7554/ eLife. 76086.
This LRT phylogeny runs contra to iconic textbook human cladograms that include Australopithecus (Fig 2) and omit Hylobates and kin. In the LRT ‘Lucy’ nests (Fig 1) among the great apes between orangutans (Pongo) and chimps (Pan) + gorillas (Gorilla, Fig 3).
Currently I’m trying to falsify this hypothesis, reaching out to the Anthropology Dept at Washington University. I’ll let you know how that goes if a meeting happens. I don’t care where the falsification comes from. Send data if you have some insight that includes gibbons AND australopithecines. Good luck on that. I’ve looked without much to show for that search.
Figure 2. The gibbon lineage leading to humans in the LRT. Australopithecus is added for comparison. The brown/black bones are restored, including the unknown feet.
Today let’s cherry pick several additional traits, both morphological and behavioral, that link gibbons to humans exclusive of the other, more robust apes (Fig 3). I’ll bring in some published experts, Ankel-Simons 2007 and Fleagle 2013.
Figure 3. Ape (hominoid) skeletons to scale. Gibbons, other than their arms, have human-like proportions and a similar upright bipedal stance. Note the relatively large braincase relative to the small mandible and small face in the gibbon and human.
Ankel-Simons 2007 wrote: “At first glance, one can see that the facial part of the skull of the lesser apes [= gibbons] is comparatively small, and the overall shape of the skull is dominated by the large braincase.
See figure 4. Also notable is the presence of eyebrows, lacking in chimps.
Figure 4. Homo longi (aka ‘dragon man’) shown with the distored face of Hylobates applied. Those are eye brows, absent in chimps, surmounting the ridges. Note the relatively flat face, protruding in chimps. Note the wide set eyes, wider than the lips, not wider in chimps.
Ankel-Simons 2007 continued: “In addition, the orbital openings of both the lesser and great apes are comparatively large and typically larger than in Old World monkeys. In species of both Hylobates and Symphalangus [= gibbons], the supraorbital tori, also known as brow ridges, that are so characteristic of great apes are not expressed.”
“The mandible is comparatively slender and shallow in lesser apes [= gibbons], and the ascending branches or rami of those mandibles do not rise very high.
See figure 6 for skull comparisons of Homo sapiens, Homo erectus and Hylobates. If this is indeed a phylogenetic series the skull gets longer with H erectus and taller with H sapiens.
Figure 5. Two gibbons, one running, one walking bipedally. Two color phases, one blonde, the other gray. A third color, black, is also common.
Fleagle 2013 wrote: “The gibbons, from Southeast Asia are the smallest, the most specifically diverse, and the most numerically successful of living apes [except humans]. The numerous gibbon species are relatively uniform in morphology. All are relatively small (5–11 kg), with no sexual size dimorphism.
They have simple molar teeth characterized by low, rounded cusps and broad basins. Their incisors are relatively short, but broad.
The canines are longer in gibbons, shorter in Ardipithecus, Oreopithecus and Homo. Terrestrial humans have a gender size dimorphism, distinct from arboreal Hylobates.
Figure 6. Skulls of Hylobates, Ardipithecus, Homo florensis, Homo erectus and Homo sapiens to a common brow to tooth row scale. The skull becomes longer in Homo erectus, taller with Homo sapiens.
Figure 6a. The ape line of primates here including ‘Homo’ naledi and Australopithecus, two bipedal chimps, convergent with the gibbon-human line.
Fleagle 2013 continued: “Gibbons have short snouts and shallow faces, large orbits with protruding rims, and a wide interorbital distance. Their braincase is globular and has no nuchal cresting. Only occasionally do they develop a sagittal crest. The mandible is shallow and has a broad ascending ramus.
Gibbons are outstanding among living primates in their limb proportions. They have the longest forelimbs relative to body size of any living primates, and they also have very long legs. They have long, curved, slender digits on their hands and feet as well as a long muscular pollex and hallux.
“Gibbons are the only apes that consistently have ischial callosities [they sit on tree limbs], and females also show small sexual swellings that change shape and color during the estrous cycle. [Other primates display much larger sexual swellings. Humans reduce this to zero]. They use a wide variety of both seated and suspensory feeding postures”.
Figure 7 Female gibbon callosities and minimal sexual swelling. This is distinct from quadrupedal chimps and monkeys, more like bipedal humans. Humans lost callosities when they stopped sitting on tree branches high in the canopy.
Fleagle 2013 continued: “Gibbons specialize on a diet of ripe fruit, part of which is found in small, widely scattered clumps throughout the forest and part of which occurs in large bonanzas, such as many figs. Gibbon species also eat varying amounts of new leaves and invertebrates, such as termites and arachnids.
Humans are omnivores and cook food for faster digestion.
“Most gibbons live in small groups composed of one male, one female, and up to four dependent offspring.
As do humans.
However, some populations may have more than one individual of either sex. Larger groups are particularly common in some siamang populations on Sumatra. In addition, extra-pair matings have been reported for many gibbon populations.
As do humans.
Numerous hypotheses have been put forth to explain pair-bonding in gibbons, including paternal care, mate-guarding, resource defense, and infanticide defense.
Pair bonding is distinct from bonobos and chimps, similar to humans.
Figure 8. Gibbon mother behind bars. Note the gracile build and long legs, as in humans. Note the relatively small ear, lower than the skull, much larger in chimps, smaller in gorillas
Fleagle 2013 continued: “Gibbons have single births every four or five years, with considerable individual variance in female reproductive success. Siamang males carry their offspring during its second year of life. In other species, male investment is not so extensive.
As do humans.
Young gibbons spend up to 10 years in their family group before leaving, usually after harassment by their same-sex parent. There is some evidence that gibbons may have a very long lifespan for their size. For example, one H. muelleri lived for 60 years in captivity.
As do humans.
“All gibbons are fiercely territorial and defend their core areas with daily calling bouts and occasional intergroup conflict. Apart from vocal duetting, social interactions within a gibbon group are usually limited and consist primarily of occasional grooming bouts”.
As do humans. Is this where singing and music had its genesis
Miller 2010 reports, “In friendly greetings, corners of mouth are drawn back, revealing teeth, and tongue is sometimes protruding. In anger, mouth is opened and closed repeatedly, smacking lips and snapping teeth together. Snarling is interpreted as an intention of biting.” Sexual maturity: 9 years for female, 6.5 years for male. “The White-Handed Gibbons in particular are fussy eaters and prefer fruit over anything else, especially fruits high in sugar such as figs. Gibbons are omnivores which eat both plants and animals. Gibbons drink water, often by dipping a hand into the water or rubbing a hand on wet leaves, and then slurping up the water from their fur. They drink by licking their own fur after it has been wet”
“In the wild, White-handed Gibbons spend about 35% of their day actively foraging for food, 24% in non-foraging activities (travel, play, sentry, aggression, mating, grooming or vocalizing) and 41% being inactive”
“Inter-group interactions can be quite violent, and there is evidence that wounds incurred in territorial aggression have resulted in the death of combatants”
“White-Handed Gibbons can be kept on islands as they cannot swim and are afraid of the water. They can go down and drink water from the edge or lean over and use the water as a toilet.”
“Abnormal behaviour patterns include: excessive grooming; hair pulling; repetitive pacing, running, jumping or rocking; social withdrawal; and self mutilation. White-Handed Gibbons are a Lesser Ape and therefore very intelligent and will become bored easily and destructive if no enrichment or stimulation is provided. White-Handed Gibbons can destroy enclosure furnishings and vegetation if bored.”
“A social species that must be housed with at least one other individual.”
“The gestation period for a White-Handed Gibbon is 7 months approximately or 205 days. The oestrus cycle length is 27 days .One infant litter size. Twins are rare. Weaning occurs by age 2. Young remain with the family until sexual maturity. Nearly naked at birth. Able to vocalize soon after birth. Infants cling to mothers ventrum (stomach). Infants ingest solids at 4 months of age. Brachiation at 9 months.”
YouTube video featuring gibbons singing
Now let’s talk about Homo naledi (~300,000 years ago, Fig 9) from South Africa, described by Berger et al. 2015 as tall, skinny, pin-headed, with gibbon-like shoulders, a primitive pelvis and fully human hands and feet at the ends of long, skinny legs. The teeth are flat, as in humans. The face is angled and dished, as in apes. Homo naledi was a cave dweller and not associated with stone tools, despite a late appearance in the fossil record coeval with larger brained hominids. Berger indicates Homo naledi was likely around for 2-3 mya prior to its late appearance in the cave.
Homo naledi needs a new generic name. It is closer to Australopithecus than to the lineage of Homo. It is bipedal by convergence with Homo, by homology with Australopithecus. Don’t assume that Australopithecus was basal to humans just because it was bipedal and shared several traits. These were by convergence and have persisted due to taxon exclusion of the gibbon lineage.
Figure 9. Updated with the addition of Homo floresiensis and Ausralopithecus (Homo) naledi.
According to Wikipedia, “H. naledi anatomy indicates that, though they were capable of long distance travel with a humanlike stride and gait, they were more arboreal than other Homo, better adapted to climbing and suspensory behaviour in trees than endurance running. Tooth anatomy suggests consumption of gritty foods covered in particulates such as dust or dirt.”
The foot of Homo naledi (Fig 10) is often considered quite human-like, but is it? Distal tarsal #1 is missing and it seems to be key to the abduction/adduction of the pollex (= digit 1).
50-minute YouTube video from 2020 featuring Lee Berger talking about his Homo naledi discoveries. “It had almost gibbon-like shoulders.” “What is Homo naledi? We have no idea.”
Still wondering why gibbons and their bipedal kin have been traditionally omitted from human ancestor studies and cladograms. Let me know of any citations that include them. Gibbon omission needs to be repaired. Gibbons and their bipedal, gracile kin are competing candidates. Let’s figure this problem out together.
YouTube video featureing Steven Churchill also wondering if Australopithecus sediba and Homo naledi are convergent with the Homo lineage. Click to play.
References Ankel-Simons F 2007.Primate Anatomy (Third Edition). Berger LR et al. 2015. Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife. 4. doi:10.7554/eLife.09560. Fleagle JG 2013.Primate Adaptation and Evolution (Third Edition). Miller S 2010. Husbandry manual for White-handed gibbon Hylobates lar (Mammalia – Hylobatidae) Western Sydney Institute of TAFE, Richmond Course Name and Number: Certificate III Captive Animals – 1068 Lecturer: Graeme Phipps
Just so you know… even paleo PhDs can’t come to an agreement on their rhynchocephalian taxon list (Figs 1, 3–5). Some taxa make all the lists. Others don’t. Some taxa only appear in the LRT. The problem runs no deeper than tradition, cherry-picking and taxon exclusion. A solution is offered.
Chambi-Trowell et al. 2021 report, “Rhynchocephalians are a group of lizard-like diapsid reptiles that were very diverse during the Mesozoic but are now restricted to a single extant genus in New Zealand.
Sadly even the top paleontologists have not yet added enough taxa to their analyses to realize what the large reptile tree (LRT, 2084 taxa, subset Fig 1) revealed in 2011. The traditional clade Diapsida is diphyletic. Two unrelated clades developed this skull morphology by convergence. In the LRT the term ‘Diapsida’ is restricted to the archosauromorph diapsids (Petrolacosaurus and kin). By convergence lepidosauromorph diapsid-grade taxa, including rhynchocephalians, nest in the LRT in the clade Lepidosauria. The addition of basal reptile and pre-reptile taxa in the LRT recovers this basal split in the Reptilia if you want to check, these results.
Figure 1. Subset of the LRT focusing on Rhynchocephalia and outgroup taxa. Compare to other recent studies (Figures 3–5) where taxon exclusion and cherry-picking have led to phylogenetic chaos and underestimating the taxon list.
Figure 2. Micromenodon skull from Chambi-Trowell et al. 2021. Colors added here. This image is offered as a typical rhynchocephalian for readers unfamiliar with this clade.
Chambi-Trowell et al. 2021 continue, Recent cladistic analyses have revealed two major clades, Eusphenodontia and the more crownward Neosphenodontia, but relationships of individual taxa have remained difficult to determine because of missing data and an unrevised data matrix. Here we drastically revise the established data matrix on rhynchocephalians by reassessing, evaluating, and adding new characters and operational taxonomic units, differing from any previous analyses in our goal to consider all known rhynchocephalians.
Lofty goal, but the LRT includes trilophosaurs, rhynchosaurs and kin in this monophyletic clade. So the stated goal of Chambi-Trowell et al., ‘to consider all known rhynchocephalians’ falls short due to taxon exclusion based on tradition. You can tell that the authors (including textbook writer MJ Benton) are cherry-picking taxa because their outgroups include two random squamates, neither a basal taxon, one a derived gekko.
Lesson: Alwayslet your software tell you which taxa are outgroup and ingroup taxa from a wide gamut of candidates. Don’t follow tradition. Tradition might not be correct.
Figure 3. Rhynchocephalia cladogram from Chambi-Trowell et al. 2021. This cladogram differs from the others both in topology and taxon list. Short-bodied and large-limbed Vadasaurus is not a pleurosaur ancestor. See link below.
Figure 4. Cladogram from Sues and Schoch 2021 focusing on Rhynchocephalia. Colors added here. This cladogram differs from the others both in topology and taxon list.
Sues and Shoch 2021 also provide a rhynchocephalian cladogram “A new taxon of sphenodontian reptile, Micromenodon pitti new genus new species is described from the Upper Triassic (Carnian) Vinita Formation of the Richmond basin of the Newark Supergroup in Virginia. Phylogenetic analysis found Micromenodon pitti n. gen. n. sp. as an early-diverging sphenodontian with fully acrodont tooth implantation.
This specimen is based solely on a maxilla. The Sues and Schoch cladogram differs from the others both in topology and taxon list.
Figure 5. Rhynchocepahalian cladogram from Simoes, Caldwell and Pierce 2020. This cladogram differs from the others both in topology and taxon list.
But wait there’s more. Simoes, Caldwell and Pierce 2020 also produced a cladogram of rhynchocephalians (Fig 5) that differs from the others (Figs 1, 3, 4).
Only the LRT determined its rhynchocephalian taxon list by testing all competing candidate taxa and letting the software determine clade members. Other workers, many with PhDs, used tradition and cherry-picking rather than testing. In the LRT traditional outgroups, like trilophosaurus, azendohsaurs and rhynchosaurs, nest in the Rhynchocephalia. In the LRT traditional ingroups, like Homoeosaurus, nest in the Squamata. The differences seen in academic studies (above) are resolved by the wider gamut LRT (Fig 1) in full resolution.
Workers. Stop making traditional mistakes. Build your own wide gamut LRT so you won’t keep making the same taxonomic errors over and over with the approval of professors. editors, referees and textbooks. Academic consensus is failing here and elsewhere. Rely only on evidence. Sure you have an urge and a calling to focus microscopically, to be a specialist, an expert, but sometimes you have to step back and get a panoramic view, like the rest of us. Build your own wide gamut LRT to get that panoramic view.
References Chambi-Trowell S et al. (9 co-authors) 2021. The diversity of Triassic South American sphenodontians: a new basal form, clevosaurs, and a revision of rhynchocephalian phylogeny. Journal of Systematic Palaeontology 19(1):1–34. Sues H-D and Schoch RR 2021. A new early-diverging sphenodontian (Lepidosauria, Rhynchocephalia) from the Upper Triassic of Virginia, U.S.A. Journal of Paleontology , 95(2):344–350. online. Simoes TR, Caldwell MW and Pierce SE 2020. Sphenodontian phylogeny and the impact of model choice in Bayesian morphological clock estimates of divergence times and evolutionary rates. BMC Biology 18(191):1–30.
Kellner et al. 2022 reassess a taxon known from only a few scattered bones, Faxinalipterus (Bonaparte 2010) originally described as a basal pterosaur. Some bones were more extensively exposed for this study. Others were µCT scanned for the first time. Kellner et al no longer consider Faxinalipterusa pterosaur (contra Bonaparte 2010). Readers heard that here first online in 2013, about nine years ago.
Figure 1. Faxinalipterus matched to Scleromochlus in 2013. The former is more primitive, like Gracilisuchus, in having shorter hind limbs and more robust fore limbs. The maxilla with fenestra and fossa, plus the teeth, are a good match. In 2022 Faxinalipterus was split into two genera.
Back in 2013, with too few bones for testing in the LRT, Faxinalipterus was matched to the basal bipedal crocodylomorph, Scleromochlus (Fig 1), a taxon omitted from both papers (Bonaparte 2010, Kellner et al. 2022). Virtually every aspect of Faxinalipterus seems to be a good match, including chronological age and overall size. Faxinalipterus is more primitive in having shorter hind limbs and more robust front limbs, as in the Scleromochlus relative, Gracilisuchus (Fig 1 upper right).
The biggest problem with Faxinalipterus studies continues to be taxon exclusion. Kellner et al. follow other authors in excluding Scleromochlus (Fig. 1) and bipedal basal crocodylomorphs from their analyses. The authors also omit bipedal members of the Fenestrasauria (Peters 2000), Langobardisaurus, Cosesaurus, Sharovipteryx and Longisquama. These taxa are known from complete skeletons and they nest with pterosaurs in several analysis (Peters 2000). These, in turn, nest with lepidosaurs (Peters 2007, LRT), far from archosaurs. With these determined taxonomic exclusions Kellner et al. 2022 ignominiously enters the professionally embarrassing timeline of pterosaur origin studies.
Kellner et al. discuss a humerus and scapulocoracoid (MCZ 10152) McCabe and Nesbitt 2021 assigned to Lagerpeton. Earlier the possibility of a connection with Procompsognathus(Fig 2) was considered. It’s an excellent match. Even so, always beware of chimaeras. Previous authors omitted Procompsognathus from their studies.
Figure 2. Procompsognathus had proportions that precisely fit the MCZ 101542 material shown here as a color photo.
From the Kellner et al. abstract: “Our interpretations on the identity of several bones differ from those of the original description, and we found no support favoring pterosaur affinities for the taxon.
Confirmation of the 2013 hypothesis. Good to hear.
“The maxilla previously referred to Faxinalipterus minimus is disassociated from this taxon and referred to a new putative pterosauromorph described here from a partial skull and fragmentary postcranial elements. Maehary bonapartei gen. et sp. nov. comes from the same fossiliferous site that yielded Faxinalipterus minimus, but the lack of overlapping bones hampers comparisons between the two taxa.
At times like this, a third, more complete taxon, with parts that overlap both enigma taxa, can resolve such issues. Unfortunately, Kellner et al. 2022 omitted a suitable third taxon, Scleromochlus (Fig 1) despite this solution appearing online for the last 9 years.
As longtime readers realize, and the pterosaur origin timeline documents, this is how pterosaur workers operate. They omit without testing and thereby keep their blinders on. Each one seeks their own hopeful headlines while ignoring competing citations.
“Our phylogenetic analysis places Faxinalipterus minimus within Lagerpetidae and Maehary bonapartei gen. et sp. nov. as the earliest-diverging member of Pterosauromorpha.”
Their phylogenetic analysis cherry-picks taxa and omits actual pterosaur ancestors, which Peters 2000 labeled Fenestrasauria. ‘Pterosauromorpha’ is a junior synonym for Reptilia in the LRT which tests all competing candidates. Faxinalipterus is related to the crocodylomorph archosaur, Scleromochlus, not to the pterosaur lepidosaurs.
Figure 3. Skull elements from Faxinalipterus transferred to Maehary by Kellner et al. 2022. The authors did not realize the palate bone was a taphonomically jumbled collection (bottom grayscale image) here digiitally segregated (middle colored image). Compare this reconstructed palate to the one in Scleromochlus.
Have you noticed the emerging geographic patterns in pterosaur origin studies? The English prefer their own Scleromochlus to the exclusion of lagerpetids. South Americans prefer their own lagerpetids to the exclusion of Scleromochlus.
Colleagues. please stop wasting your time trying to re-discover the ancestry of pterosaurs by cobbling together bits and pieces of unrelated taxa. Please stop omitting the complete skeletons that have already been shown to be ancestral to pterosaurs (Peters 2000) just because you can, and your editor David Hone, will let you. Please stop hoping/pretending that Lagerpeton and kin are close to pterosaurs and dinosaurs. Adding taxa moves Lagerpeton close to the proterochampsid Tropidosuchus (Novas and Agnolin 2016) as confirmed by South American workers here.
Every one of these silly attempts at finding pterosaur ancestors where you want them, instead of where they are, just makes the paleo profession look less scientific and more desperate for attention while maintaining a smokescreen of elitism. No matter how you feel about ‘the messenger’ don’t omit the taxa. Go study them. They exist.
References Bonaparte JF, Schultz CL and Soares MB 2010. Pterosauria from the Late Triassic of southern Brazil. In S. Bandyopadhyay (ed.), New Aspects of Mesozoic Biodiversity, Lecture Notes in Earth Sciences 132:63-71. Kellner AWA, Holgado B, Grillo O, Pretto FA, Kerber L, Pinheiro FL, Soares MB, Schultz CL, Lopes RT, Araújo O, Müller RT 2022. Reassessment of Faxinalipterus minimus, a purported Triassic pterosaur from southern Brazil with the description of a new taxon. PeerJ 10:e13276 http://doi.org/10.7717/peerj.13276 McCabe MB and Nesbitt SJ 2021. The first pectoral and forelimb material assigned to the lagerpetid Lagerpeton chanarensis (Archosauria: Dinosauromorpha) from the upper portion of the Chañares Formation, Late Triassic. Palaeodiversity, 14(1) : 121-131. Novas FE and Agnolin FL 2016Lagerpeton chanarensis Romer (Archosauriformes): A derived proterochampsian from the middle Triassic of NW Argentina. Simposio. From Eventos del Mesozoico temprano en la evolución de los dinosaurios”. Programa VCLAPV. Conferencia plenaria: Hidrodinámica y modo de vida de los primeros vertebrados. Héctor Botella (Universitat de València, España) 2016 Peters D 2000b. A Redescription of Four Prolacertiform Genera and Implications for Pterosaur Phylogenesis. Rivista Italiana di Paleontologia e Stratigrafia 106 (3): 293–336. Peters D 2007. The origin and radiation of the Pterosauria. In D. Hone ed. Flugsaurier. The Wellnhofer pterosaur meeting, 2007, Munich, Germany p 27.
The Royal Tyrrell Museum Speaker Series recently featured Dr. P David Polly of Indiana University (video link below). He reported on the history and current status of the Circle Cliffs and Grand Staircase-Escalante national monuments in Utah. This is a very interesting and informative talk, well worth seeing.
Royal Tyrrell Museum Speaker Series on YouTube. Click to play. Approximately 1 hour 10 minutes in length.
One image from Dr. Polly’s online video talk caught my eye. But what really grabbed my attention was the headline Dr. Polly gave to the image: “Ancestor of crocodiles and dinosaurs?” (Fig 1). Polly reported this specimen was ‘under study’.
I shouldn’t have to say this, but discoveries like this should not be shown in public prior to publication. Publication online constitutes a form of publication.
All that aside, let’s focus on Dr. Polly’s headline (Fig 1). He’s teasing the idea of linking crocs to dinos.
Figure 1. Image from Royal Tyrrell Museum Speaker Series featuring P David Polly. He reports this skull was found in the Staircase-Escalante region of Utah and is ‘under study’. Yellow highlight box added here.
Traditionally only the large reptile tree (LRT, 2084 taxa) recovers a topology in which crocodiles and dinosaurs had a last common ancestor: the basalmost archosaur PVL 4597 (Figs 2, 3).
Given that backstory, now you know why Dr. Polly’s headline (Fig 1) came as an unexpected surprise. It indicates that finally, at least some workers are wondering about or recovering the hypothesis of croc + dino interrelationships (Figs 2, 3) first advanced here ten+ years ago and cemented since then with more and more pertinent taxa.
Did the LRT help? Or was this convergence? It doesn’t matter.
Figure 2. Subset of the LRT focusing on the base of the Archosauria. That’s where we find the last common ancestor of crocodiles dinosaurs in the LRT that Dr. Polly’s caption in figure 1 wonders about.
I ran the data on Dr. Polly’s photo and am able to answer Dr. Polly’s question, but since the specimen is ‘under study’, let’s put a pin in that. Meanwhile, here (Figs 2, 3) are some old PterosaurHeresies images that detail and illustrate the topology of the basal archosaur subset of the LRT in which crocs and dinos have nested as sister clades virtually from the beginning.
Figure 3. The origin of dinosaurs and crocodiles in the LRT to scale. Gray arrows show the direction of evolution. This image includes Decuriasuchus, Turfanosuchus, Gracilisuchus, Lewisuchus, Pseudhesperosuchus, Trialestes, Herrerasaurus, Tawa and Eoraptor. Note the phylogenetic miniaturization at the origin of the clade Archosauria (Crocs + Dinos).
If and when any paper comes out that links crocodiles to dinosaurs, remember then, you heard it here first.
References Get back to this spot when that paper is published.
These two storks (or stork-like kingfishers) (Figs 1, 2) are identical, but geographical separated cousins with inflatable necks. The South American jabiru(Jabiru mycteria, Lichtenstein 1819) has been described as ‘the largest and most striking of the American storks.’
Figure 1. Skeleton and in vivo views of Jabiru.
By contrast, the skeletally identical African marabou (Fig 2, Leptoptilos crumenifer, Lesson 1831), was described by the same author as ‘amongst the most hideous birds in the world’, ‘ungainly’ and ‘grotesque’.
Figure 2. Skeleton and living marabou ‘stork’ Leptoptilos.
These two taxa would have been congeneric if found in the fossil record. Geography has separated these two over a hundred million years ago, coeval with many famous dinosaurs, then terror birds and finally primitive peoples.
Figure 3. Skull of Jabiru. Compare to Leptoptilos in figure 4.
Figure 4. Skull of Leptoptilos. Compare to Jabiru in figure 3.
These two nearly congeneric birds now living on opposite sides of the Atlantic Ocean add evidence to a common origin in the Early Cretaceous. That hypothesis was first explored here with similarly split hornbills and toucans (Fig 5).
Figure 5. South America and Africa during the Albian, 100 mya. This is when toucans and hornbills must have separated.
With basalmost crown birds appearing in the Early Cretaceous and derived birds, like the marabou, jabiru, hornbill and toucan radiating before the Atlantic had become an ocean, and penguins appearing in the Paleocene, more and more birds, maybe most of them, seem to have had their genesis and radiation in the Cretaceous.
Figure 6. Subset of the LRT focusing on marabous and their relatives. Here long-legged stork-like taxa appear to be derived. Terns and kingfishers are basal taxa. Sea gulls and plovers are outgroup taxa.
As predicted yesterday additions to the LRT are going to start looking more and more like current members. The geographic separation of these two sisters (Figs 1, 2) is the real story here. As this example demonstrates, sometimes the key part of the story is not always found in the skeleton.
References Lesson R 1831. Traité d’Ornithologie Lichtenstein H 1819. Archives of Natural History, Volume 35(1):88-99. Linnaeus C 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata.
The number of taxa currently present in the large reptile tree (LRT, 2082 taxa), large pterosaur tree (LPT, 261 taxa) and therapsid skull tree (TST, 78 taxa) are at levels in which phylogenetic gaps have faded. That means all taxa nesting close to one another are strongly similar in morphology, size, niche and diet. That indicates these three cladograms strongly model actual evolutionary events. That also predicts discoveries to come will likely resemble currently tested taxa. Exceptions are always welcome.
That also means the three tree topologies have all reached numbers in which their curves are leveling off, reaching their plateau.
Plateau definition: a part of a graph in which the value of the ordinate shows little or no change with increasing value of the abscissa. Alternately: a period of (relative) stability in the course of a progression.
This growing online project started in earnest 12 year ago Adding taxa weekly and making corrections (=’housekeeping‘) transparently is what made this project unique.
The LRT, LPT and TST are not and will never be perfectly scored. The data came from too many sources. Some data will always remain incomplete, lost during taphonomy. Even so, these three cladograms are each large enough to test smaller more focused studies for taxon exclusion. Even so, corrections will continue as long as mistakes and omissions appear. If you see any, let me know.
Looking to the future Suggestions for new taxa are always welcome, but here the search for new taxa has slowed to a crawl. We knew this day would eventually arrive. It’s been a great run punctuated by novel hypotheses seemingly appearing weekly. As others announce new taxa and new interrelationships some will be added to the cladograms. Others will be reported and checked for problems, as usual. In any case, the pace will slow. The puzzle is more or less complete.
These five problems continue to plague paleontology: Number one: taxon exclusion. Number two: borrowing untested cladograms. Number three: trusting genomic results. Number four: trusting textbooks and academic traditions. Number five: freehand reconstructions. Solution: Keep adding taxa to your own trait-based cladogram. Trace specimens with transparent colors and from those tracings create more accurate and verifiable reconstructions.
Figure 1. From the Beginning – The Story of Human Evolution (Little Brown 1991) available as a FREE online PDF at DavidPetersStudio.com
At that time no one had any idea that whales, turtles and even diapsids were diphyletic. That pterosaurs arose from cosesaurs and were bipedal lepidosaurs. That the first amniotes were amphibian-like gephyrostegids. That only crocodylomorphs and dinosaurs were archorsaurs. That sharks arose from sturgeons. That bony fish arose from hybodontid sharks. That at least ten of the 13 Solnhofen birds were different enough to nest apart from one another. That bats have living ancestors. Etc. etc.
At that time only a few intrepid workers were experimenting with the onerous drudgery of using software and digital matrices in an attempt at keeping phylogenetic tradition and bias to a minimum. But there was a hack for that.
Unfortunately, many paleo myths persist. So the work is not over. But the work can coast now, at least until the next issues arise.
Thank you for your readership, even if you held on waiting for a big fail.
Thank you for your readership, even when you knew you had to do it secretly so your colleagues wouldn’t find out.
Thank you for your readership especially when you saw through the errors presented here and out there.
Most of us have matured and learned over the past decade. For some, the whining continues. For others, the testing and experimenting continues. For the last decade the LRT carried the wide gamut cladogram baton. For the next few decades brilliant younger workers are needed to carry that same baton. Specialists will always be produced by universities. Occasionally a generalist is needed to resolve broader issues.
I hope others will replicate this online experiment for refutation, confirmation or that gray area, correction, so it won’t be unique. I hope someone is already assembling their matrix. If not, see what you can do to encourage someone or a group of someones who love vertebrate evolution to produce a competing candidate cladogram based on traits (not genes). Otherwise this hypothesis of interrelationships will forever remain a singular untested hypothesis. Taxon exclusion will rule.
This is your invitation to take your favorite science, vertebrate paleontology, to the next level.
Just wanted to take a moment to remember Dr. Hal Levin, professor emeritus, Earth and Planetary Sciences, Washington University, St. Louis, USA. He entered our profession in 1961, became department chair in 1973 and died in 2019 at age 90. He was the author and often illustrator for eleven editions of his geology textbook, “The Earth Through Time.”
Fig 1. Hal Levin, Wash U professor and author
From Levin’s obituary: “Throughout his life, Levin was committed to unraveling the Earth’s secrets by examining microscopic ancient invertebrates from the Earth’s sedimentary layers to predict future changes and to expose the deleterious effects of climate change.
“Levin joined the Washington University faculty in 1961. He taught and conducted research while concurrently serving as the coordinator of preprofessional studies in Arts & Sciences.
“He was chair of the Department of Earth and Planetary Sciences from 1973-76. A prolific author, Levin wrote and illustrated numerous textbooks and laboratory studies in geology and paleontology, including 11 editions of the popular geology textbook “The Earth Through Time.”
Back in the day when I was illustrating books like ‘Giants‘ …and back when you could just walk into university libraries and offices without a digital ID and a face mask, Dr. Levin invited me to illustrate the cover for the fifth edition of his university-level textbook on geology, ‘The Earth Through Time’ (Fig 2). I was just getting started in paleontology back then. He was just getting ready for retirement.
Figure 2. ‘The Earth Through Time’ the fifth of eleven editions, by Harold (Hal)( Levin, WashU. Cover illustration by David Peters.
Wash U is not famous for fossils, but has several impressive legacy specimens from the old Gustav Hambach collection. Habach was a German immigrant who became head of the geology dept in 1887. Since then Wash U has been more into geology, both here on Earth and on Mars, with forays into human origins during the heyday of David Tab Rasmussen and Glenn C Conroy.
Figure 3. Ichthyosaur from the Hambach collection, Washington U, St Louis, USA.
Hal Levin was always kind and accommodating, perhaps because there have never been more than a few paleo professionals and enthusiasts in St. Louis. Somehow we all find our mentors.