A large, scrappy and disarticulated pterosaur find from Morocco, Barbaridactylus (Fig. 1), appears center stage in the newest Prehistoric Planet episode called ‘Flamboyant Flyers‘. Unfortunately, traditional myths and problems abound.
Figure 1. At bottom, the KJ2 specimen of Nyctosaurus with crest, to scale with the UNSM 93000 specimen of Nyctosaurus, to scale with the remains of Barbaridactylus and a frame from ‘Flamboyant Flyers’ showing the combination of two pterosaurs to make one imagined pterosaur for Prehistoric Planet.
Problem #1 In ‘Flamboyant Flyers’ male pterosaurs are said to be larger and decorated with cranial crests, distinct from smaller, plainer females.
Fact #1 Phylogenetic analysis shows smaller, plainer specimens are more primitive. Only derived taxa had the time to evolve ornate crests. No male/female traits have ever been supported. Bennett 1992 wondered if a large-basin pelvis represented a female Pteranodon, but did not realize that all Nyctosaurus specimens have a large-basin pelvis. Some rivaled Pteranodon in size (Fig 1).
Figure 2. David Attenborough alongside an imaginary pterosaur skull attributed to Barbaridactylus, a specimen found without a skull, documented in figure 1. The crest is too thick. Pterosaur experts should know this.
Problem #2 In ‘Flamboyant Flyers’ the giant crest is shown to be half as thick as the skull (Fig 2).
Fact #2. In pteranodons and nyctosaurs the crest is typically 1/8-inch (3mm) thick. The imagined skull shown with narrator David Attenborough comes from the model shop, not from the Early Cretaceous sands of Morocco. Only small parts of the rear mandible were found from the head of Barbaridactylus (Fig 1).
Figure 3. Frame one: Blueprint-type image form ‘Flamboyant Flyers’ showing the incorrect deep-chord, bat-wing model for pterosaur wings that has never been found in the fossil record. Frame two: Nyctosaurus images that show the correct shallow-chord wing membrane found in all pterosaur fossils that preserve soft tissue.
Problem #3 In ‘Flamboyant Flyers’ the pterosaurs wings extend to the ankles (Figs 3, 4).
Fact #3 No pterosaurs fossils have ever been found with wing membranes extending to the ankles. I keep looking for one exception that shows this morphology. There are no exceptions. Uropatagia trail each hind limb, missing from the ‘Flamboyant Flyers’ blueprint in figure 3.
Figure 4. Frame from ‘Flamboyant Flyers’ showing the traditional but incorrect pterosaur wing membrane extending to the ankles. Frame two shows the correct morphology for all pterosaur specimens that preserve soft tissue wing membranes, and thus presumed for nyctosaurs. Note the large size of the male with a distinct crest. We have no evidence for larger or more decorative males.
Problem #4 In ‘Flamboyant Flyers’ the pterosaurs are shown to be awkward quadrupeds on land. They over-bend the normally large, stiff wrist far beyond the breaking point in order to make this happen.
Fact #4 Most pterosaur tracks are quadrupedal and these are all attributed to beachcombing clades, not nyctosaurs. A minority of tracks are bipedal, some germanodactylids, basal to nyctosaurs. Due to their long metatarsals nyctosaurs are presumed to be bipeds, balancing over their toes (Fig 1), using their long forelimbs as ski poles when walking. Extended so far anteriorly, the long forelimbs of nyctosaurs can contribute no anterior vector to locomotion.
Figure 5. Barbaridactylus shown taking off by catapulting off the forelimbs, unlike birds or bats and not preserved in the fossil track record. Since pterosaurs could stand bipedally (see figure 4) they could open their wings and start flapping before becoming airborne, not wait until they were set to crash with their wings still below their bodies, creating no thrust or lift. Also note the extreme bending required at the wrist, which is impossible based on examination of the carpals in many pterosaurs.
Problem #5 In ‘Flamboyant Flyers’ the pterosaurs take to the air by bending their forelimbs way past their physical limits, unlike bats and birds.
Fact #5 Pterosaurs were bipeds (Fig 1) when they wanted to be and took to the air by flapping their large wings, like birds do from branches, water and land. No extreme wrist bending or dangerous leaping without the wings already extended and producing thrust is necessary.
Figure 6. Hatchling Barbaridactylus from Prehistoric Planet.
Problem #6 In ‘Flamyboyant Flyers’ the hatchling pterosaurs have a short rostrum and other ‘cute’ features found in mammals and archosaurs (birds + crocodilians).
Fact #6 Several embryo and hatchling pterosaurs have proportions identical to adults. None have a short rostrum, unless the adult also had a short rostrum. This isometric growth strategy is typical of lepidosaurs,, which is where pterosaurs nest in the large reptile tree. Pterosaurs also hatch from leathery eggs, typical of lepidosaurs. They also have elongate manual digit 4 and pedal digit 5, traits absent in archosaurs. We’ve known this since Peters 2000 and 2007, but university students are still taught that pterosaurs are archosaurs.
We looked at Alcione and Barbaridactylus earlier here in 2018. Barbaridactylus is twice the size of most nyctosaurs (Fig 1). Alcione had very short metacarpals, odd for a nyctosaur, unless it was flightless, which is also odd for a nyctosaur.
Figure 7. Darren Naish, lead scientific consultant for Prehistoric Planet. He did a great job on the dinosaurs.
Figure 8. Liz Martin-Silverstone, paleontologist from U of Bristol, home of MJ Benton.
Figure 9. Quetzalcoatlus sunning itself, distorted somewhat, according to the skeleton standing beside it.The myth of the bat-wing pterosaur is on display here. So is a wild imagination.
We looked at Prehistoric Planet episodes earlier here and here.
References Bennett SC 1992. Sexual dimorphism of Pteranodon and other pterosaurs, with comments on cranial crests. Journal of Vertebrate Paleontology 12: 422–434. Longrich NR, Martill DM and Andres B 2018. Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary. PLoS Biol 16(3): e2001663. https://doi.org/10.1371/journal.pbio.2001663 Peters D 1995. Wing shape in pterosaurs. Nature 374, 315-316. Peters D 2002. A New Model for the Evolution of the Pterosaur Wing – with a twist. – Historical Biology 15: 277–301. Peters D 2000b. A Redescription of Four Prolacertiform Genera and Implications for Pterosaur Phylogenesis. Rivista Italiana di Paleontologia e Stratigrafia 106 (3): 293–336. Peters D 2007. The origin and radiation of the Pterosauria. In D. Hone ed. Flugsaurier. The Wellnhofer pterosaur meeting, 2007, Munich, Germany. p. 27.
Zhou et al. 2022 found an Early Cretaceous pterosaur partial wing (Fig. 1) and a tiny complete foot (SDUST-V1006, Figs 2–4) they decided not to name yet. They reported, “The new pterosaur specimen is characterised by an unusual pedal configuration of a short and spread metatarsus with elongate digits, showing a close resemblance to the dentulous Ornithocheiroidea.”
Figure 1. The little SDUST V1006 pterosaur wing shown close to full scale @ 72dpi. Colors added here.
Zhou et al. reported, “SDUST-V1006 is distinct from all known ornithocheiroids in the pedal proportions, possibly representing a derived condition of the digital elongation. Furthermore, its pedal configuration that the pedal digits are extremely longer than the relevant metatarsals is also distinct from other pterosaur clades.”
Figure 2. Fengning pterosaur right foot, greatly enlarged. Photo and diagram from Zhou et al. 2022. Colors added here. Pedal digit 5 was lost and left a faint mark on the surface.
The Zhou et al. linear-plot diagram of the second wing phalanx and metatarsal 3 in pterosaurs nested the SDUST specimen close to Haopterus and Boreopterus, two basal ornithocheirids. Their phylogenetic analysis nested the SDUST specimen basal to Istiodactylidae + (Boreopteridae + Ornithocheirae). As it turned out, this was by convergence, something linear-plot diagrams are unable to understand given that only two traits are tested (= Pulling a Larry Martin).
Here, in a competing phylogenetic analysis both with and without pedal digit 5, the Fengning pterosaur (SDUST-V1006) nests between Wukongopterus and Kunpengopterus (Fig 4) in the large pterosaur tree (LPT, 261 taxa).
Figure 4. Mddle Jurassic Kunpengopterus compared to scale with SDUST-V1006, an Early Cretaceous pterosaur with much larger wings and a smaller pes.
With present data the Fengning pterosaur (SDUST-V1006) is the first known wukongopterid to survive into the Early Cretaceous. It is convergent with the clade of Late Jurassic scaphognathids that evolved to become Early Cretaceous ornithocheirids. Shifting the Fengnng pterosaur to Boreopterus in the LPT adds 8 steps based on foot traits only. So that’s not trivial. Testing all the available traits, not just a few, even if just a few are available for testing, still remains the best way to identify enigma taxa.
Zhou et al concluded, “The first pterosaur is described from the Huajiying Formation, the lowest horizon of the Early Cretaceous Jehol Biota, in the northern Hebei Province, China. It is tentatively assigned as an indeterminate ornithocheiroid, showing an unusual pedal configuration with short and spread metatarsus and elongate digits. This pedal configuration is in contrary to the general pattern of the elongate metatarsus and shorter digits in archaeopterodactyloids and pteranodontians, as a derived condition like that of boreopterids and istiodactylids. Functionally, the elongate digits might offset the shortened metatarsus to enlarge the pedal surface for paddling, representing a new strategy in adaptation to the aquatic environment.”
With such tiny feet (Fig. 4), paddling seems unlikely, even with interdigital webbing.
References Zhou C-F, Zhu Z and Chen J 2022. First pterosaur from the Early Cretaceous Huajiying Formation of the Jehol Biota, northern Hebei Province, China: insights on the pedal diversity of Pterodactyloidea. Historical Biology, DOI:10.1080/08912963.2022.2079085
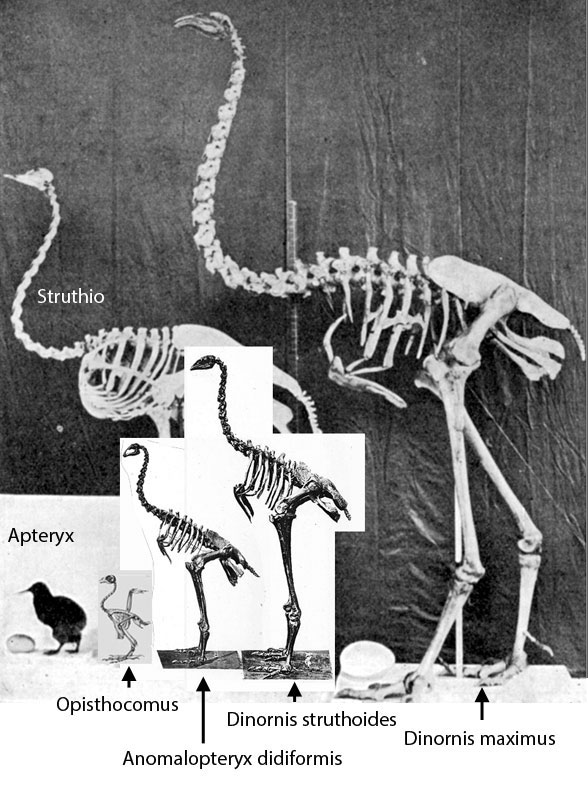
Figure 1. Anomalopteryx compared to Dinornis and Opisthocomus.
Like Dinornis, Anomalopteryx lacked forelimbs (= wings). The neck was proportionally longer in larger taxa.
Figure 2. Anomamalopteryx skeletons. Note the loss of the pectoral girdle and pedal digit 1.
Nearly ever trait is identical in these two New Zealand flightless birds, which means, maybe they are congeneric, as Owen 1843 originally named this taxon.
Figure 3. Skull of Anomalopteryx in three views. Colors added here.
Anomalopteryx didiformis(originally Dinornis didiformis Owen 1844, Rechenbach 1853) is the recently extinct bush moa of New Zealand. It was about the size of a turkey and was the smallest known moa. Note the reduction of the antorbital fenestra to a vestige here and in the largest moa, Dinornis.
Figure 4. Gondwana at 83mya. Arrows locate the present locations known for hoatzins (Opisthocomus) in the Amazon, and related moas (Dinornis) in New Zealand. Guineafowl live in sub-Saharan Africa today.
Members of this wide-ranging and ancient clade are chiefly terrestrial (= none swim, dive or soar over seas), seed eaters and rapid, burst flappers that do not migrate. They range in morphology from as plain as a brown sparrow, to as spectacular as a peacock and include the tallest of all birds, Dinornis (Fig 1) from New Zealand, most closely related to the odd South American hoatzin, Opisthocomus. Their last common ancestor had to live before Zealandia split from Antarctica and South America about 100mya (Fig 4).
Figure 5. Subset of the LRT focusing on birds.
References Owen R 1843. On the remains of Dinornis, an extinct gigantic struthious bird. Proceedings of the Zoological Society of London: 8–10, 144–146. Rechenbach L 1853. Avium Systema Naturale.
Updated June 12, 2022 and Augut 13, 2022 with the moving of most placoderms to a more basal position close to Ordovician jawless fish and developing jaws convergently with basal gnathostomes like Chondrosteus.
Li et al. 2021 reported they were describing, “a new Silurian fish close to the common ancestor of modern gnathostomes.”
That topology is not supported by the LRT (subsets Figs 4, 5) where placoderms develop jaws by convergence with modern gnathostomes (represented by toothless Chondrosteus, Fig 6) and its outgroup taxa, sturgeons and ostracoderms.
Placoderm ancestors are derived from Arandaspis in the LRT (Figs 4, 5).
The LRT solved the long-standing ‘origin of jaws’ problem (Fig 6) a few years ago by simply adding taxa. On June 2022 a second origin of jaws was documented.
Figure 1. Bianchangichthys, a tiny placoderm. Colors added here based on tetrapod homologies. Also shown full scale at lower right. This is the original published reconstruction. Compare to updated reconstruction using DGS added September 4, 2023.
Figure 1b. Bianchangichthys under DGS. Colors and reconstruction added here September 4, 2023.
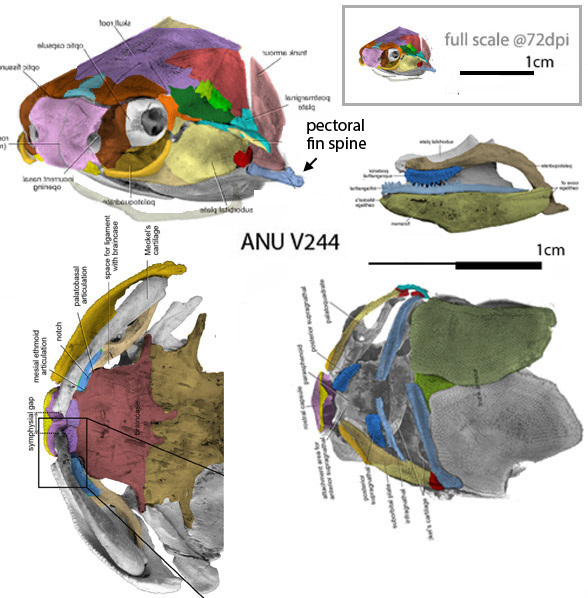
Taxon exclusion mars the Li et al. description of their really tiny placoderm, Bianchengichthys (Fig 1). The large reptile tree (LRT, 2096 taxa then 2110 taxa on June 12, 2022) nests Bianchenichthys with the ANU V244 specimen (Fig 2). These two tiny taxa nest at the base of the clade that ultimately produced mid-sized Coccosteus (Fig 3) and giant Dunkleosteus. So, this is yet another example of phylogenetic miniaturization (Fig 3) at the genesis of new clades recovered by the LRT.
Bianchengichthys micros Li et al. 2021, Latest Silurian, IVPP V27128A and B) is one of the smallest and most basal arthrodire placoderms. The oribit + naris is anteriorly oriented. If the lacrimal and postorbital are present they are extremely tiny. Bianchengichthys was reported to be, “exquisitely preserved with mandible and paired fins. Systematically, the new taxon is close to the last common ancestor of bony and cartilaginous fishes. The mandible interestingly combines characters from arthordires and Entelognathus.”
Figure 2. The unnamed tiny Late Silurian transitional placoderm, ANU V244 was preserved in perfect 3d. Colors added here.
From the Li et al. 2021 abstract: “The Silurian Period occupies a pivotal stage in the unfolding of key evolutionary events,including the rise of jawed vertebrates.“
An ancestor of Chondrosteus (Fig 6) is the last common ancestor of all gnathostomes (= jawed vertebrates) in the LRT.
The earliest known gnathostome is Early Silurian, Loganiella, a small basal shark.
“However, the understanding of this early diversification is often hampered by the patchynature of the Silurian fossil record, with the articulated specimens of jawed vertebrates onlyknown in isolated localities, most notably Qujing, Yunnan, China.”
The LRT is not ‘patchy’. The LRT includes such a wide gamut of extinct and extant taxa that patches and gaps are gone. In this way ‘this early diversification” of any included clade is well represented and clearly documented (Figs 4, 5) back to pre-Cambrian worms.
The Li et al. abstract continues: “Here, we report a new Silurian maxillate placoderm, Bianchengichthys micros, from the Ludlow of Chongqing, with a near-complete dermatoskeleton preserved in articulation.”
See figure 1.
“Although geographically separated, the new taxon resembles the previously reported Qilinyu in possessing a unique combination of dermatoskeletal characters.”
Qilinyu nests close to Biachengichthys in the LRT (Fig 4).
“However, the dermal bone of the mandible in Bianchengichthys unexpectedly differs from those in both Qilinyu and Entelognathus and displays a broad oral lamina carrying a line of tooth-like denticles, in addition to the marginal toothless flange.”
Be careful with expectations (= biases). Just score your taxa and let your software tell you what interrelationships develop. Bianchengichthys may represent a second, third or fourth appearance of jaws, depending on the definition. In placoderms derived from jawless arandaspids the change from jawless to jawed (Fig 3) appears to be spectral representing a transitional zone with different experiments in ‘jaws’ present‚ not a firm line.
The abstract continues: “The external morphology of the pectoral fin is preserved and reveals an extensively scale-covered lobate part, flanked by a fringe of lepidotrichia-like aligned scales.“
The preservation of fins in placoderms is rare. But note ancestral Eurynotus (Fig 3) preserves hard and sharp fins.
“The phylogenetic analysis reveals that Bianchengichthys is positioned immediately below Entelognathus plus modern gnathostomes.”
Taxon exclusion is the problem. Adding taxa changes the Li et al. tree topology (Figs 4,5). Keep adding taxa until your tree topology stops changing.
“The discovery significantly widens the distribution of Silurian placoderm-grade gnathostomes in South China and provides a range of morphological disparity for the outgroup comparison to the earliest evolution of jaws, dentitions, and pectoral fins in modern gnathostomes.”
No. You can’t make those conclusions with such small, cherry-picked cladograms. This is especially so when there is a large online cladogram (the LRT) that presents a more panoramic topology that reveals the mistakes of smaller studies. Colleagues: add taxa! Repair your cladograms.
“We also demonstrate that the previously reported Silurian placoderms from centralVietnam are maxillate placoderms close to Qilinyu, Silurolepis, and Bianchengichthys,corroborating the paleogeographic proximity between the Indochina and South Chinablocks during the Middle Paleozoic.”
The Viet Nam placoderm(s) are represented by scraps.
Figure 6. Origin of jaws from the ostracoderm, Hemicyclaspis, Thelodus, Acipenser (sturgeon) and Chondrosteus.
Fellow workers, please add taxa before presenting your next cladogram. Your specimens and discoveries deserve more attention to the phylogenetic setting you put them in. Get it right. Don’t take the easy way out. Keep adding taxa until your topology no longer changes every time you add taxa.
References Hu Y, Lu J and Young GC 2017. New findings in a 400 million-year-old Devonian placoderm shed light on jaw structure and function in basal gnathostomes. Nature Scientific Reports 7: 7813 DOI:10.1038/s41598-017-07674-y Li Q et al. 2021. A new Silurian fish close to the common ancestor of modern gnathostomes. Current Biology 31:3613–3620. https://doi.org/10.1016/j.cub.2021.05.053
Figure 1. Homo longi, the dragon man, compared to Hylogbates. colors and overlays added here. Hylobates distorted to fit Homo longi. Note the beard, eyebrows, flat narrow face not found in Australopithecus and Pan, the chimpanzee.
Here the skull of Homo longi is overlaid with an appropriately distorted face of Hylobates to imagine a flesh and fur layer over the nearly human skull. Of course this purposefully overlooks the nudity and development of sweat glands of humans. Note the presence in Hylobates of a human-like beard and eyebrows, but still no mustache. These traits are not found in the long-faced, knuckle-walking chimp and gorilla lineage that produced a convergent biped, Australopithecus, and probably ‘Homo’ naledi (Fig 3).
Figure 2. The bipedal gibbon lineage leading to humans alongside the convergent Australopitecine (gray background).
Speaking of Homo naledi the pes (= foot) is traditionally reconstructed as human-like (Fig 3), but distal tarsal #1 is missing. The pollex was possibly abducted, oriented in a more chimp-like fashion (Fig 3). Much depends on that missing distal carpal (Fig 4).
Figure 3. Several ape pedes, including ‘Homo’ naledi. Frame 2 abducts the pollex medially to more closely resemble ape feet. Note the lineup of the proximal metatarsals in ‘Homo’ naledi a in the three other apes. This is distinct from the inset of metatarsal 2 in humans, Oreopithecus and barely in Hylobates in figure 4.
Also note the small detail where metatarsal 2 meets tarsal 2. In humans, Oreopithecus and barely in Hylobates (Fig 4), this joint is offset from the others due to the small size of tarsal 2. This makes the hinge here less flexible, better for walking. By contrast, in Australopithecus, ‘Homo’ naledi (Fig 3) and apes this tarsal 2 joint is not offset. It is aligned with the others, creating a flexible hinge at this part of the foot, better for climbing.
Figure 4. Various hominoid pedes compared. Colors added here. Note the misaligned metatarsal 2 due to a short carpal 2 in humans, Oreopithecus and barely in Hylobates. This is distinct from the metarsal 2 and distal tarsal 2 in Australopithecus, apes and ‘Homo’ naledi in figure 3.
Video of several speakers (listed below) describing their heretical insights into the origin of humans.
YouTube video on “Origins of Genus Homo” Several heretical hypotheses are expressed in this video. Several align with hypotheses presented here.
Australopithecus sediba was possibly convergent as a biped @ 5:24 by speaker Steven Churchill. The ability to run bipedally (shared with gibbons), but attributed only to Homo @ 28:00 by speaker Daniel Lieberman. Ranging, running @42:00 by speaker Herman Pontzer. Side note: The gracile gibbon lineage (Fig 2) is the most wide ranging lineage (Africa, Europe, China, SE Asia). Primates burn half the calories for a mammal of the same size @49:45 Humans share food. Most other mammals and primates do not share. Gibbons do share.
Russell 1975 wrote about FMNH PR 846: “By far the finest specimen of Globidens known. The specimen consists of a nearly complete skull and anteriormost 11 presacral vertebrae.”
From Russell 1975, three views of the FMNH PR 846Globidens skull. Colors added here.
Globidens dakotensis (Gilmore 1912, Russell 1975, Late Cretaceous, Pierre Shale, skull 66cm long) nests with Plotosaurus in the LRT. Note the truncated epipterygoids not reaching the jugals, the contact between the prefrontal and the fused postfrontal + postorbital. The vomer is either taphonomically absent or absent in vivo. The nasals are absent. The jugal dips below the orbit. The teeth are sphere-shaped. The pterygoids do not reach the quadrates. The lacrimal is fused to the maxilla. Many of these traits are distinct from other tested mosasaurs.
References Russell DA 1975. A new species of Globidens from South Dakota, and a review of globidentine mosasaurs. Fieldiana Geology, 33(13): 235-256. wiki/Globidens
YouTube video from Prehistoric Planet. Click to play
Another clip from Prehistoric Planet follows traditional thinking regarding Mononykus (Figs 1–3) in imagining this alvarezsaurid as a termite eater. Narrator, David Attenborough, says Mononykus (‘one claw’) means “giant claw”. The animation is excellent, just a little misguided.
Figure 1. Clip from the Prehistoric Planet video. Frame 2 compares the animated version to the skeleton of Mononykus. Frame 2 compares it to Shuvuuia.
The animators gave their dinosaur an imaginary woodpecker-like tongue for extracting termites from rotten logs. As you can see (Fig 1) the skull is unknown in leggy Mononykus and the overall proportions are different. Shuvuuia is also shown, all three to the same metatarsal length. Maybe ‘Mononykus’ was easier for Attenborough to say.
Figure 2. Giant Deinocheirus, a contemporary of Mononykus, might have served as the host and dining room for a series of ever smaller and more specialized parasite eaters.
The Prehistoric Planet animators are doing an excellent job. Their science advisor is Darren Naish.
Figure 3. Forelimb of Mononykus. Large deltopectoral crest pulls humerus toward the sternum like a clasp. The claws permanently point medially. Not ventrally.
Mononykusolecranus (Perle et al, 1993) Late Cretaceous ~70 mya, 1 m in length was a flightless alvarezsaurid theropod. Only digit I remained on the hand, which was otherwise stunted, except for the proximal ulna (the elbow) which was enlarged. The hindlimbs were elongated. The tibia had two cnemial crests and the femur/tibia joint was very flexible. Metatarsal 3 did not reach the tarsus.
Shuvuuia deserti (Chiappe, Norell and Clark 1998, Late Cretaceous; IGM 100/1304) is a smaller alvarezsaurid that retains vestigial digits 2 and 3 and a longer tail. This alvarezsaurid pattern is characterized by the strong, but stunted forelimbs likely used to cling to feathery and scaly hosts as insect-eating ‘tick-birds’ to larger dinosaurs. (IGM 100-0977 is a larger specimen)
YouTube video from Prehistoric Planet. Click to play
Deinocheirus does make an appearance in Prehistoric Planet and the plot line indicates it is beset with biting insects! Where did that idea come from?? Unfortunately, the science advisor did not provide the poor giant dinosaur with tiny tick-bird-like companions clinging to its feathers to eat the pests and provide some relief. No, the science advisor provided a nearby, rather mundane dead tree for Deinocheirus to rub up against. Opportunity lost.
References Chiappe LM, Norell MA and Clark JM 1998. The skull of a relative of the stem-group bird Mononykus. Nature, 392(6673): 275-278. Choiniere JN, Xu X, Clark JM, Forster CA, Guo Y, Han F 2010. A basal alvarezsauroid theropod from the Early Late Jurassic of Xinjiang, China. Science 327 (5965): 571–574. Perle A, Norell MA, Chiappe LM and Clark JM 1993. Flightless bird from the Cretaceous of Mongolia. Nature 362:623-626. Perle A, Chiappe LM, Rinchen B, Clark JM and Norell 1994. Skeletal Morphology of Mononykus olecranus (Theropoda: Avialae) from the Late Cretaceous of Mongolia. American Museum Novitates 3105:1-29.
Turnix suscitator (Gray 1840, Figs 1, 2) is the extant buttonquail, a poor flyer that looks like and behaves like a quail. Beyond that, everyone else has had an opinion, a hypothesis or a genomic analysis that considered Turnix a member of three other clades.
Figure 1. Turnix, the button quail, is a basal pigeon in the LRT.
According to Wikipedia, “buttonquail are small, drab, running birds that avoid flying. Buttonquail were traditionally placed in Gruiformes or Galliformes (the crane and pheasant orders).”
By contrast, based on genomic testing (Prum 2015), Turnix is an outgroup to Glareola, which is an outgroup to Uria. None of these interrelationships are recovered by the large reptile tree (LRT, 2092 taxa). With every added taxon results from Prum 2015 continue to fare no better than alchemy.
Also by contrast, the LRT nests this Turnix with basal pigeons likePterocles, the extant chestnut-bellied sandgrouse.As we learned earlier, the LRT nested Glareola as a pre-nightjar. Uriawas nested a pre-penguin.
Figure 2. Turnix skull. Colors added here. Note the absence of the postfrontal and the squamosal that does not extend laterally beyond the quadrate. The ectopterygoid (amber) reappears here along an anteriorly extended pterygoid (dull red).
The Turnix skull has more bones than usual and fewer bones than usual. The postfrontal and postorbital are absent. The squamosal does not extend lateral to the quadrate. A newly returned (= reversal) slender ectopterygoid extends to the maxilla. Haven’t seen those traits yet in related taxa.
This appears to be a novel hypothesis of interrelationships. If not, please provide a citation so I can promote it here. It shouldn’t be novel. These interrelationships should have been established years ago, but famous bird workers (like Prum 2015) started leaning on genes and lost their way.
Figure 3. Subset of the LRT focusing on birds.
References Gray GR 1840. List of Gen. B:59 Prum et al. (6 co-authors) 2015. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 526:569–573. online
The holotype of the pachypleurosaurHanosaurus (IVPP V 3231, Young 1972, Fig 1) is exposed in dorsal view, recovered in several parts.
Figure 1. The holotype of Hanosaurus is exposed in dorsal view, the opposite of the referred specimen in figure 2.
The new referred specimen assigned to Hanosaurus (IVPP V 15911, Wang et al. 2022, Fig 2) is preserved in ventral view and is fully articulated. In the large reptile tree (LRT, 2092 taxa) these two nest together matching all traits possible given their opposite exposures.
Figure 1. The IVPP V 15911 specimen referred to Hanosaurus, confirmed by the LRT.
Problems arise when Wang et al presented their sauropterygian cladogram (Fig 2; 62 taxa, 181 characters) which suffers from taxon exclusion, including labeling the outgroup as ‘ancestor’. The text reveals they cherry-picked Youngina capensis (AMNH 5661) as that outgroup ‘ancestor’ taxon.
Which Youngina? The Youngina capensis holotype (AMNH 5561) is not basal to the Enaliosauria (= Sauropterygia + Ichthyosauria) in the LRT, but nests basal to terrestrial protorosaurs and archosauriforms. However the referred BPI 3859 specimen incorrectly referred to Youngina capensis nests with Orovenator and is basal to terrestrial and aquatic younginiforms in the LRT (Fig 3). Specimen numbers were not provided by Wang et al.
Other taxa preceding the resurrected clade Enaliosauria (= Sauropterygia + Ichthyosauria) are also missing from Wang et al. (Figs 2–4).
Figure 2. Frame 1: Cladogram from Wang et al. 2022. Frame 2: Color overlay indicates clades in the LRT. Blue/Cyan = Ichthyopterygia. Violet/Lavendar = Thalattosauria. Pink = Eosauropterygia. Yellow/green = Placodontia. This Wang et al. cladogram could use more ingroup and outgroup taxa. The ‘ancestor’ is Youngina capensis. Compare to a subset of the LRT in figure 3.
There is a continuing trend displayed in Wang et al. 2022 for academics to show vague outlines of taxa (Fig 2) instead of documenting their scoring with DGS (color) overlays of insitu specimens that can be used to create understandable and comparable reconstructions, as in ReptileEvolution.com. Don’t take the easy way out, even if it is acceptable to your colleagues and editors. Make your methods transparent. Don’t use occasional generic silhouettes (Fig 2).
Figure 3. Subset of the LRT focusing on basal Enaliosauria and marine younginiiforms.
Hanosaurus hupehensis (Young 1972, Early Triassic 249 Ma, IVPP V 3231) is small pachypleurosaur from the Middle/Late Triassic of China, known from scattered parts. Originally it was described as a thalattosaur.
Figure 4. Subset of the LRT reduced to the Wang et al taxon list. Compare to figures 2 and 3.
Rieppel (1998) nested Hanosaurus between Keichousaurus and Anarosaurus, but the LRT nests it in the Keichousaurus clade (Figs 3, 4) with or without excluded taxa.
References Riepppel O 1998. THe systematic status of Hanosaurus hupehensis (Reptilia, Sauropterygia) from the Triassic of China. Journal of Vertebrate Paleontology 18(3):545-557. Wang W, Shang Q, Cheng L, Wu X-C and Li C 2022. Ancestral body plan and adaptive radiation of sauropterygian marine reptiles. bioRxiv preprint doi: https://doi.org/10.1101/2022.04.25.489368 Young CC 1972. A marine lizard fron Nanchung, Hupeh province. Memoirs of the Institute of Vertebrate Paleontology and Paleoanthropology. Academia Sinica 9:17-28.
According to SciAm.com “Steve Brusatte is a professor at the University of Edinburgh and author of The Rise and Reign of the Mammals, a new narrative history of mammal evolution (Mariner Books, 2022).”
Brusatte is a good writer, full of superlatives, active verbs and exciting adjectives. He’s popular with the public and the media. Unfortunately, Brusatte keeps demonstrating his lack of understanding the subject matter. He’s young. He hasn’t done the necessary analyses. He repeats what he learned in school. Too often his own words have internal conflicts (see below).
Sometimes young scientists need to curb their impulses to write books and use that time to create their own wide gamut cladogram in order to confirm and establish interrelationships. Then, with that authority, is the time to write the book. You don’t want to spread misinformation in permanent media, especially if that includes the potential for decades of professional embarrassment due to printed errors.
Figure 1. Ectoconus is an Early Paleocene, sheep-sized terrestrial former tree shrew.
Brusatte begins with the discovery of a new Ectoconus fossil: “This fossil mammal, Ectoconus, was a revolutionary. It lived a mere 380,000 years after the worst day in Earth history, when a six-mile-wide asteroid ended the Age of Dinosaurs in fire and fury, ushering in a new world.”
First time we’ve ever heardEctoconuswas “a revolutionary.” Brusatte doesn’t report how it was a revolutionary, other than it was a placental mammal, like us. It’s not a new taxon. Cope described a complete skeleton (Fig 1) in 1884.
Brusatte writes, “We actually know very little about the mammals that endured the extinction and persevered during the next 10 million years, during the Paleocene epoch.”
We actually know quite a bit about “the mammals that endured the extinction”, according to Brusatte himself! After all he “actually” wrote an entire book about it (see above), and the rest of the SciAm article (see below). This is example #1 of internal conflict mentioned above. We’ve seen this many times before. Paleontologists like to set up imagined problems so they can come in as heroes with imagined solution, too often failing due to taxon exclusion.
“Dinosaurs became giants and excluded mammals from large-bodied niches. Mammals did the opposite: with their small body sizes, they could exploit ecological niches that the bigger dinosaurs couldn’t access.”
Readers: Is this correct? Giant vs small? Or is this over simplification? Were there no small to medium dinosaurs? No medium to large mammals, like… Ectoconus (Fig 1)? [Internal conflict #2]. Maybe the situation is more complex than “dinosaurs became giants”.
“A bounty of pint-sized mammals—none larger than a badger—lived underfoot of the dinosaurs.”
“One such group—the multituberculates—flourished in the Cretaceous underworld… Meanwhile, as multituberculates prospered, three other groups quietly branched off on their own. These trailblazers gave rise to the three mammal lineages that persist today.”
After analysis multituberculates are highly derived members of placental clde Glires, the gnawing clade. We have Jurassic porcupines (Maiopatagium). Brusatte doesn’t know this.
“These trailblazers“(= multituberculates) did not give rise to the three mammal lineages that persist today. That’s a textbook myth Brusatte repeats without testing.
“The molecular clock—a technique that uses DNA differences among modern species and back calculates to estimate when they diverged—predicts that some placental lineages, including primates, lived alongside the dinosaurs. Although paleontologists are desperate to recover fossils of such early placentals, they have yet to be found.”
The LRT uses traits to determine the same conclusion. Look at Nasua, the coatimundi, and the very similar lemur, Lemur. Both are basal placentals, likely appearing in the Early Jurassic, and still living today! This phylogenetic information was overlooked by Brusatte. DNA (= genomic testing) is to be avoided in deep time analyses.
Figure 2. The coatimundi (Nasua) compared to the ring-tailed lemur (Lemur).
Brusatte wrote, “Perhaps surprisingly, mammals were doing well in the latest Cretaceous. At least 30 species lived in Montana back then, filling many ecological roles at the base of the dinosaur-dominated food chain, including bone crunchers, flower eaters, insectivores and omnivores.”
Bone crunchers? Flower eaters? Could Brusatte be more specific? His readers are not children. Here I’m going to guess: Bone crunchers are triisodontids = archaic hippos crunching grasses and pond plants. Flower eaters are herbivores in general, from rabbits and squirrels to deer and pygmy opossums since ‘herbivores’ do not make Brusatte’s list even though his Ectoconus (Fig 1) was an herbivore.
“The vast majority of these creatures were metatherians (early members of the marsupial line) or multituberculates. Early cousins of placentals called eutherians were present, though rare.”
Early cousins of placentals called eutherians?The LRT does not recover this distinction. Nor does Brusatte explain this distinction. Earlier he said there there were only three mammal lineages, which is correct. Brusatte makes it sound like there was one more.
“The earliest Paleocene scene is dire. There is a fossil locality in Montana dated to approximately 25,000 years after the asteroid hit, called the Z-Line Quarry. It reeks of death. Almost all the mammals that flourished in the region in the Cretaceous are gone; only seven species remain.”
Seven species? Which seven? How diverse or restricted?
“Several other fossil sites divulge what was happening over the next 100,000 to 200,000 years. If you pool together all mammals from this time, there are 23 species.”
Let’s take a moment to digest this. Brusatte is telling his readers how extremely rare fossils are in general. He tells us what all paleontologists know. There are only a few tiny windows of time within a few tiny windows of geography that preserve fossils. That means there are huge expanses of unaccounted, unsurveyed, undug time and geography that multiply those numbers by simple extrapolation.
Brusatte reports, “A paltry 7 percent of mammals survived the carnage.”
Earlier Brusatte reported 7 of 30 species survived, then 23 of 30 species. Neither of those represent seven percent. Or is Brusatte talking raw numbers from two tiny windows in time and space. In any case, which ones? How? The word “How” is in the headline, so we’re looking for a payoff.
Brusatte provides only this clue, “This bleak state of affairs raises a question: What allowed some mammals to endure? The answer became apparent when Wilson Mantilla looked at the victims and survivors. The survivors were smaller than most of the Cretaceous mammals, and their teeth indicate they had generalist, omnivorous diets. The victims, on the other hand, were larger, with more specialized carnivorous or herbivorous diets.”
Remember Brusatte’s parameters:“The survivors were smalller” and omnivores. The “victims… were larger” and herbivores. These parameters can be immediately tested by the larger herbivorous Early Paleocene survivor Ectoconus (Fig 1) featured by Brusatte in his introduction. This is internal conflict #3.
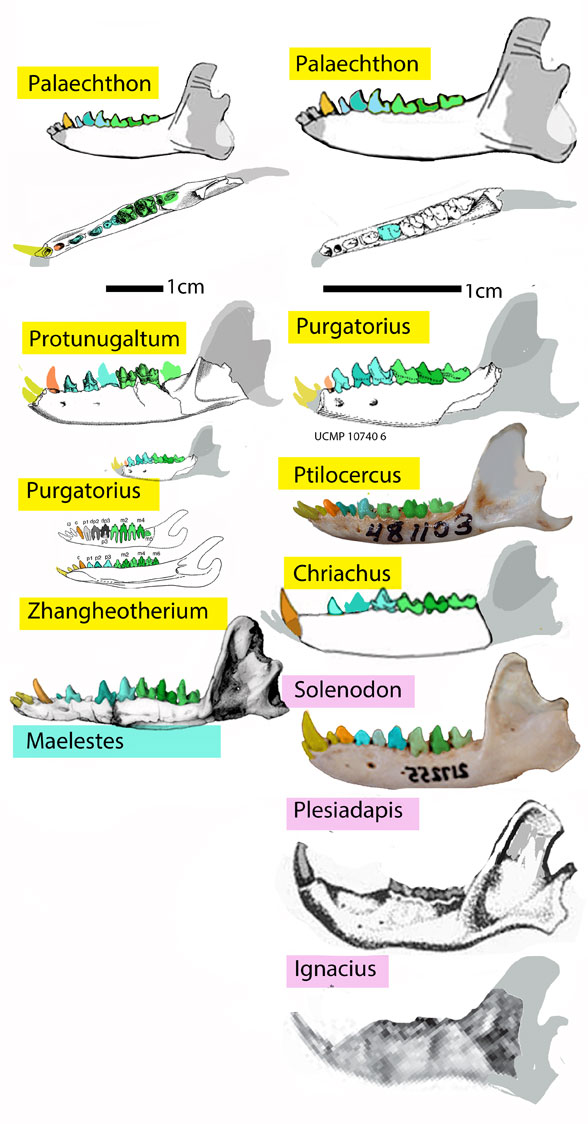
Brusatte writes: “Around 100,000 years postasteroid a new eutherian appeared in Montana and swiftly became common. Purgatorius, with gentle molar cusps for eating fruits and highly mobile ankles for clinging and climbing in the trees, was an early member of the primate line.”
This is incorrect. Purgatorius (Fig 3) is a late survivor of a Jurassic radiation of shrinking colugos, like Palaechthon. Basalmost primates are rather larger lemurs, similar in size and shape to basalmost carnivores, like living coatimundis and raccoons, also Jurassic in origin.
Figure 3. Purgatorius compared to other basal and often Paleocene mammals.
Brusatte wrote, “Ectoconus, whose skeleton we excavated in 2014, was one of these trailblazers. When it was bounding through swampy rain forests and snacking on leaves and beans 65.6 million years ago, it was the largest mammal that had ever lived there.”
Internal conflict #3 repeated. This is just the sort of placental Brusatte said did not make it through the asteroid hit (see above). Since this specimen was found in New Mexico, not far from the Yucatan impact, Brusatte needs to explain this anomaly. This is the ‘how’ we’re wondering about, but Brusatte never gets around to explaining.
Brusatte wrote, “The Paleocene mammals were not so easily categorized, however. They were clearly much larger than any Cretaceous mammals”
Here Brusatte reports the survivors were “larger than any Cretaceous mammals”. Earlier Brusatte reported, “The survivors were smaller than most of the Cretaceous mammals.“ Someone is not proofreading Brusatte’s text for internal conflict.
“and they lacked epipubis bones at the front of their pelvis, suggesting they had large placentas to nourish their young in utero. Thus, they were assuredly placentals. But their skeletons seemed peculiar—stocky and muscle-bound, with mashups of features seen in various groups of modern-day mammals.”
I realize Brusatte is writing for a popular audience, but epipubes are absent in some marsupials, present in some placentals. Putting all your phylogenetic faith in one or a few traits is called “Pulling a Larry Martin“. Only a last common ancestor determines which fossil taxa are in one clade or another.
Brusatte wrote, “Tom and his crew have collected thousands of fossils, which paint a vivid picture of Paleocene life within the first million years of the asteroid.”
Notice: the picture has gone from ‘bleak‘ to ‘vivid‘, both describing only two small windows of fossil localities. Neither is close to a million years after the asteroid. Or is there a third locality?
Figure 4. A subset of the LRT representing the second half of placental evolution. Ectoconus nests with the pantodonts in the right column.
Bursatte continues: “Among the roster of archaic placentals are animals like Ectoconus, which are shoehorned into a nebulous group called condylarths.”
Shoehorned? Does that mean it’s not a good fit? Why does Bursatte even mention ‘Condylarthra‘ when Wikipedia indicates this term has little to no academic standing and a traditional nebulous meaning?
“Members of this group were mainly plant eaters or omnivores with sturdy builds; many of them had hooves. They shared the herbivore niches with pantodonts—barrel-chested leaf gobblers with enormous hands and feet, which achieved sizes comparable to modern cows.”
In the LRT Ectoconus is precisely a pantodont, the clade basal to living edentates. That’s a phylogenetic story completely missed by Brusatte. And once again, were the survivors smaller or larger than their Cretaceous ancestors? This time they are comparable to modern cows.
Figure 5. Traditional Taeniodonta in a cladogram. With more taxa this clade splits up according to the colors shown here.
Brusatte continues: “Another group, the taeniodonts, were gargoyle-esque diggers, which used their huge clawed forearms to tear through dirt and their massive jaws and enlarged canines to root out tubers.”
Traditional taeniodonts are paraphyletic (Fig 5). So this is not a clade. Mistakes like these keep undercutting Brusatte’s authority.
“All these mammals would have feared the triisodontids, the terrors of the Paleocene, which looked like wolves on steroids and smashed the bones of their prey with crushing molars.”
As explained yesterday, triisodontids were archaic hippos, herbivores with crushing molars, not ‘wolves on steroids’. Early carnivores were like coatimundis, raccoons and meerkats.
“We are building a vast data set of fossil and extant mammals, and their anatomical and genetic features, so that we can construct a master family tree.”
That’s a great concept! More taxa create a better “master family tree” (subset Fig 4).
“Our preliminary results are encouraging. Some of the archaic species, such as taeniodonts, might have stemmed from Cretaceous eutherian ancestors and thus would be among the most primitive placentals on the trunk of the family tree.”
Keep adding taxa, Dr. Brusatte until you find taeniodonts are not a natural group and ‘eutherians’ are placentals going back to the Jurassic.
“Others, including some of the condylarths, share features with today’s hoofed mammals and are probably proto-horses and proto-cattle.”
Probably? Yes, they are proto-cattle (Fig 4). That’s what you find after testing.
“Although the precise locations of condylarths and taeniodonts and their archaic ilk in the family tree remain to be worked out, we are already grasping what they were like as living, breathing animals.”
The precise locations have already been worked out in the LRT with full resolution. Next efforts from anywhere will either confirm, refute or correct the LRT.
Brusatte writes, “Chief among these hallmarks of placental mammals is the capacity to birth well-developed young, which gestate for a prolonged period inside the mother before being born in an advanced state.”
Actually not so well developed in the first half of placental evolution, when many placental babies remained small, helpless and nest-bound, like humans, mice and dogs. In the second half (Fig 4) placentals are much better developed, able to walk behind, swim with or cling to their mothers almost immediately after birth.
Brusatte reports, “Larger [placental] offspring could more easily grow into larger adults, which may have enabled the first placentals to rapidly balloon in stature within a few hundred thousand years of the dinosaurs’ demise, after 160 million years of being stuck at tiny sizes.”
This sounds like another Brusatte generalization with plenty of exceptions. For instance, wombats and kangaroos are big marsupials while bats and shrews are small placentals.
Figure 6. A large marsupial Diprotodon museum mount and dorsal views of the manus and pes.
Brusatte reports, “The relative brain sizes of the archaic placentals were laughably small compared with not only those of today’s mammals but even those of the Cretaceous species living with the dinosaurs. The first placentals, it seems, got so big so fast that their brains couldn’t initially keep pace.”
This sounds like another Brusatte generalization with plenty of exceptions. Which Late Cretaceous placentals? Which Early Paleocene placentals? Were they related? If Brusatte could show this happening in a lineage, that would support his claim. Such a lineage is known (Fig 7), and it includes Ectoconus. Unfortunately lacking his own LRT, Brusatte is not aware of this natural clade. Again, do the work, THEN write the book and the article.
Figure 7. The origin of edentates includes the large, clawed herbivorous Paleocene Ectoconus, an asteroid survivor from New Mexico. How it survived is not explained by Brusatte 2022.
In summary, Brusatte doesn’t tell his SciAm readers “how mammals conquered the world”. That would have been fascinating. The ‘how’ remains eagerly awaited. Instead Brusatte repeats old tropes and untested hypotheses, ignores internal inconsistencies and demonstrates a need to get up to speed phylogenetically. No doubt Dr. Brusatte has a full daily agenda at this stage in his young life: writing books, traveling to dig sites and conferences, his own scientific studies, working with students, perhaps raising a family. A busy day makes it difficult to put in as much time as needed (several years) to get up to speed phylogenetically.
References Brusatte S 2022.How mammals conquered the world after the asteroid apocalypse. ScientificAmerican.comarticle Cope ED 1884. The Amblypoda. The American Naturalist 18 (112):6=461-471. Shelley SL, Williamson TE and Brusatte SL 2015. Resolving the higher-level phylogenetic relationships of “Triisodontidae” (‘Condylarthra’) within Placentalia, October 2015, Society of Vertebrate Paleontology (abstract)