
Investigation of the Membrane Localization and Interaction of Selected Flavonoids by NMR and FTIR Spectroscopy

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. FTIR Investigation of DPPC Liposomes at 25 °C
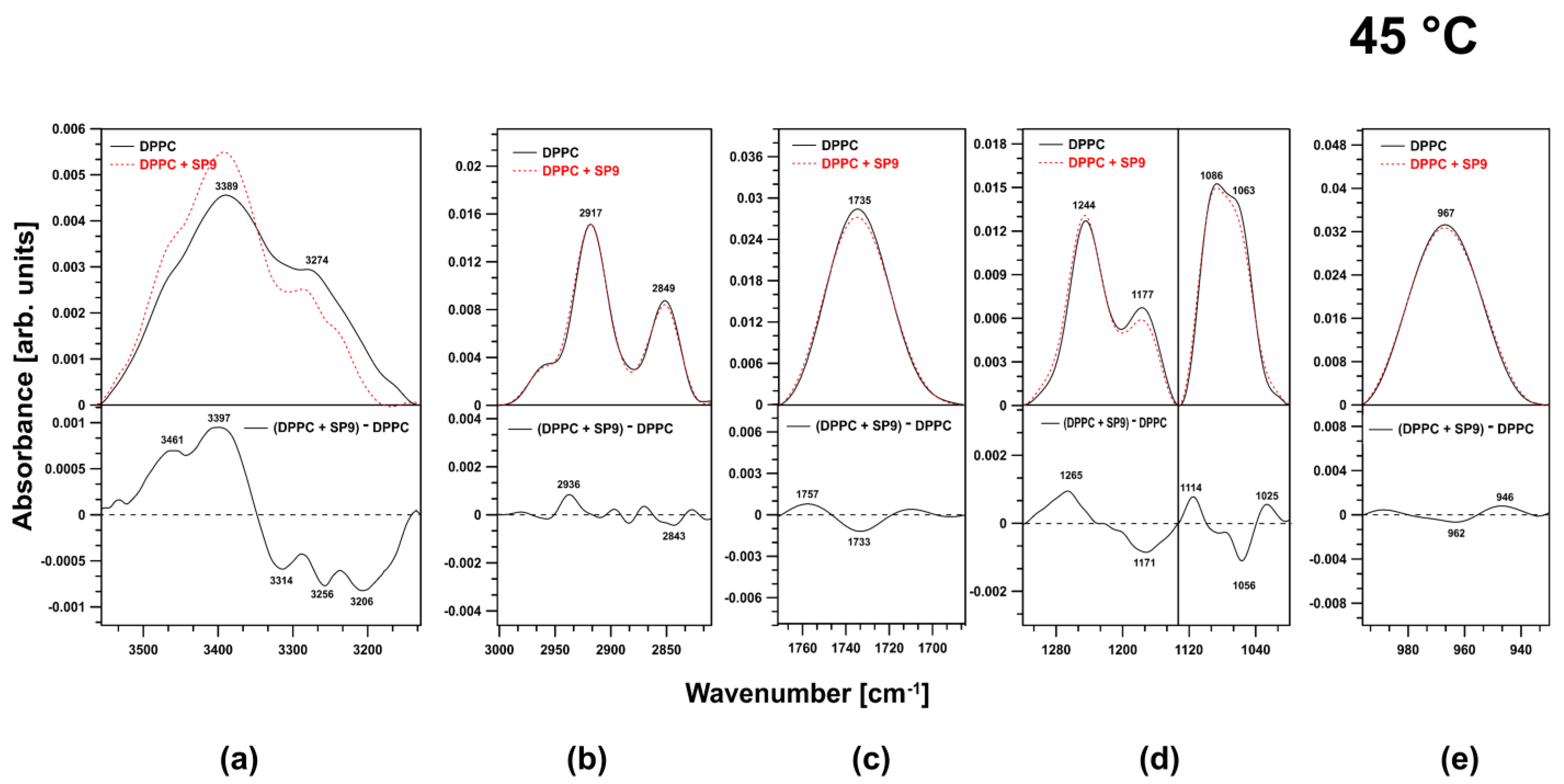
2.2. FTIR Experiments on DPPC Membranes at 45 °C
2.3. FTIR Investigation of the EYPC Membranes
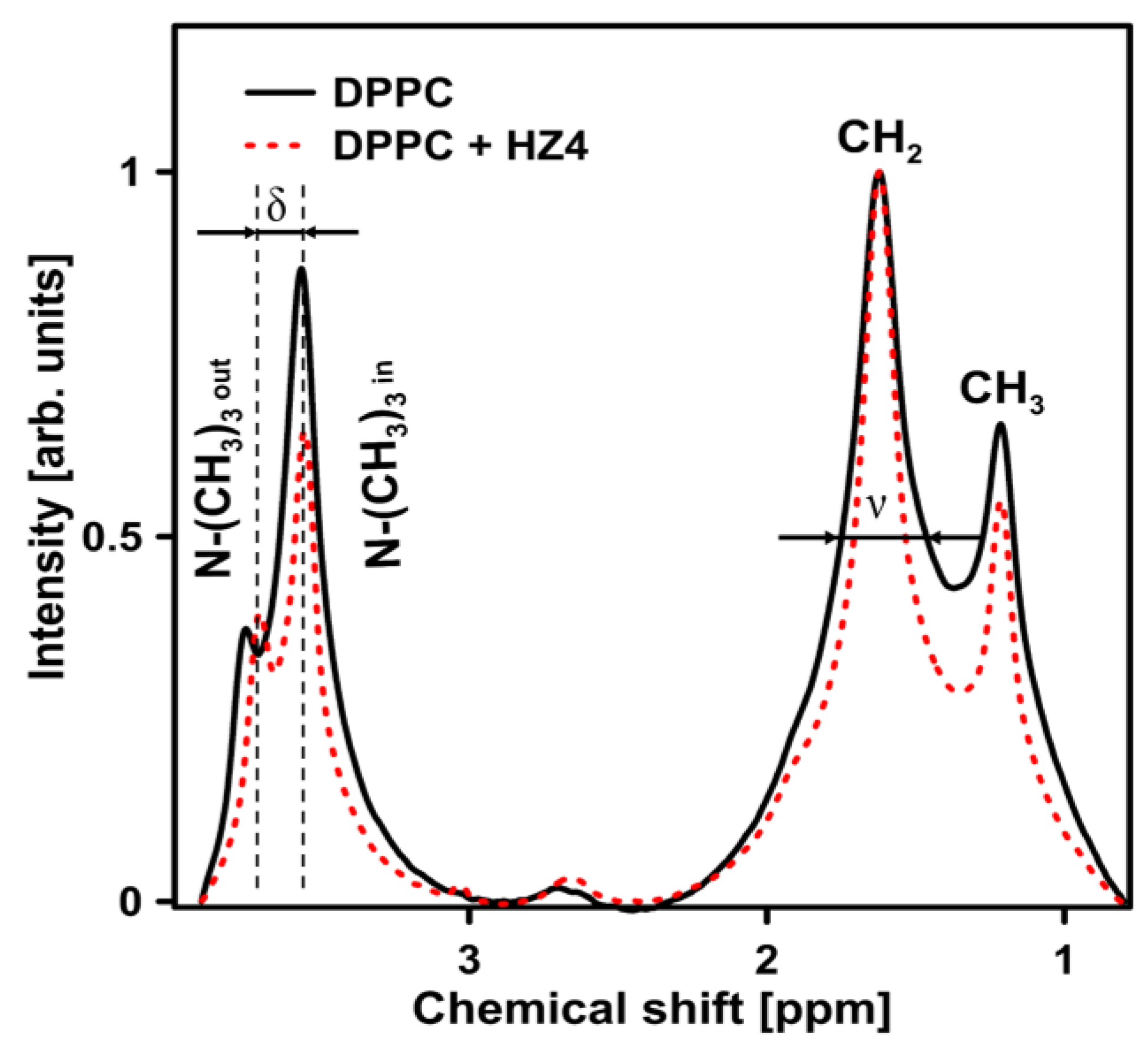
2.4. NMR Experiments on the DPPC Liposomes
2.5. Physicochemical Parameters of the Studied Compounds HZ4 and SP9
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Preparation of Liposomes
4.3. Nuclear Magnetic Resonance (1H NMR) Spectroscopy of DPPC Liposomes
4.4. Fourier-Transform Infrared (FTIR) Spectroscopy
4.5. Calculation of Physicochemical Parameters
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, K.H. Discovery and development of natural product-derived chemotherapeutic agents based on a medicinal chemistry approach. J. Nat. Prod. 2010, 73, 500–516. [Google Scholar] [CrossRef]
- Nasim, N.; Sandeep, I.S.; Mohanty, S. Plant-derived natural products for drug discovery: Current approaches and prospects. Nucleus 2022, 65, 399–411. [Google Scholar] [CrossRef]
- Chopra, B.; Dhingra, A.K. Natural products: A lead for drug discovery and development. Phytother Res. 2021, 35, 4660–4702. [Google Scholar] [CrossRef] [PubMed]
- Hendrich, A.B. Flavonoid-membrane interactions: Possible consequences for biological effects of some polyphenolic compounds. Acta Pharmacol. Sin. 2006, 27, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Erlejman, A.; Verstraeten, S.V.; Fraga, C.G.; Oteiza, P.G. The interaction of flavonoids with membranes: Potential determinant of flavonoid antioxidant effects. Free Radic. Res. 2004, 38, 1311–1320. [Google Scholar] [CrossRef]
- Sanchez-Galego, J.I.; Lopez-Revuelat, A.; Sardine, J.L.; Hernandez-Hernandez, A.; Sanchez-Yague, J.; Llianillo, M. Membrane cholesterol contents modify the protective effects of quercetin and rutin on integrity and cellular viability in oxidized erythrocytes. Free Radic. Biol. Med. 2010, 48, 1444–1454. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Gupta, S. Apigenin: A promising molecule for cancer prevention. Pharm. Res. 2010, 27, 962–978. [Google Scholar] [CrossRef] [PubMed]
- Rzymowska, J.; Gawron, A.; Pawlikowska-Pawlega, B.; Jakubowicz-Gil, J.; Wojcierowski, J. The effect of quercetin on induction of apoptosis. Folia Histochem. Cytobiol. 1999, 37, 125–126. [Google Scholar]
- Strawa, J.W.; Jakimiuk, K.; Pawlikowska-Pawlęga, B.; Gruszecki, W.I.; Kapral-Piotrowska, J.; Wiater, A.; Tomczyk, M. Polar localization of new flavonoids from aerial parts of Scleranthus perennis and Hottonia palustris and their modulatory action on lipid membranes properties. Biochim. Biophys. Acta Biomembr. 2023, 1865, 184142. [Google Scholar] [CrossRef]
- Tarahovsky, Y.S.; Muzafarov, E.N.; Kim, Y.A. Rafts making and rafts braking: How plant flavonoids may control membrane heterogeneity. Mol. Cell. Biochem. 2008, 314, 65–71. [Google Scholar] [CrossRef]
- Oteiza, P.I.; Erlejman, A.G.; Verstraeten, S.V.; Koen, C.L.; Fraga, C.G. Flavonoid-membrane interactions: A protective role of flavonoids at the membrane surface? Clin. Dev. Immunol. 2005, 12, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Margina, D.; Ilie, M.; Manda, G.; Neagoe, I.; Mocanu, M.; Ionescu, D.; Gradinaru, D.; Ganea, C. Quercetin and epigallocate-chin gallate effects on the cell membranes biophysical properties correlate with their antioxidant potential. Gen. Physiol. Biophys. 2012, 31, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Veiko, A.G.; Sekowski, S.; Lapshina, E.A.; Wilczewska, A.Z.; Markiewicz, K.H.; Zamaraeva, M.; Zhao, H.C.; Zavodnik, I.B. Flavonoids modulate liposomal membrane structure, regulate mitochondrial membrane permeability and prevent erythrocyte oxidative damage. Biochim. Biophys. Acta–Biomembr. 2020, 1862, 183442. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.; Byrem, T.M.; Nair, M.G.; Strasburg, G.M. Modulation of liposomal membrane fluidity by flavonoids and isoflavonoids. Arch. Biochem. Biophys. 2000, 373, 102–109. [Google Scholar] [CrossRef]
- Herrerias, T.; Oliveira, A.A.; Belem, M.L.; Oliveira, B.H.; Carnieri, E.G.S.; Cadena, S.M.S.C.; Noleto, G.R.; Martinez, G.R.; Oliviera, M.B.M.; Rocha, M.E.M. Effects of natural flavones on membrane properties and cytotoxicity of HeLa cells. Rev. Bras. Farmacogn. 2010, 20, 403–408. [Google Scholar] [CrossRef]
- Tsuchiya, H.; Nagayama, M. Garlic allyl derivatives interact with membrane lipids to modify the membrane fluidity. J. Biomed. Sci. 2008, 15, 653–660. [Google Scholar] [CrossRef]
- Tsuchiya, H.; Nagayama, M.; Tanaka, T.; Furusawa, M.; Kashimata, M.; Takeuchi, H. Membrane-rigidifying effects of anti-cancer dietary factors. BioFactors 2002, 16, 45–56. [Google Scholar] [CrossRef]
- Karonen, M. Insights into Polyphenol-Lipid Interactions: Chemical Methods, Molecular Aspects and Their Effects on Membrane Structures. Plants 2022, 11, 1809. [Google Scholar] [CrossRef]
- Uekusa, Y.; Kamihira-Ishijima, M.; Sugimoto, O.; Ishii, T.; Nakamura, S.K.; Tanji, K.; Naito, A.; Nakayama, T. Interaction of epicatechin gallate with phospholipid membranes as revealed by solid-state NMR spectroscopy. Biochim. Biophys. Acta 2011, 1808, 1654–1660. [Google Scholar] [CrossRef]
- Rothwell, J.A.; Day, A.J.; Morgan, M.R.A. Experimental determination of octanol-water partition coefficients of querecetin and related flavonoids. J. Agric. Food Chem. 2005, 53, 4355–4360. [Google Scholar] [CrossRef]
- Fraczkiewicz, R. In silico prediction of ionization. In Comprehensive Medicinal Chemistry II; Taylor, J.B., Triggle, D.J.B.T.-C.M.C.I.I., Eds.; Elsevier: Oxford, UK, 2007; pp. 603–626. [Google Scholar]
- Comer, J.; Tam, K. Lipophilicity profiles: Theory and measurement. In Pharmacokinetic Optimization in Drug Research; Verlag Helvetica Chimica Acta: Zürich, Switzerland, 2001; pp. 275–304. [Google Scholar]
- Scheidt, H.A.; Pampel, A.; Nissler, L.; Gebhard, R.; Huster, D. Investigation of the membrane localization and distribution of flavonoids by high-resolution magic angle spinning NMR spectroscopy. Biochim. Biophys. Acta 2004, 1663, 97–107. [Google Scholar] [CrossRef]
- Tsuchiya, H. Structure-dependent membrane interaction of flavonoids associated with their bioactivity. Food Chem. 2010, 120, 1089–1096. [Google Scholar] [CrossRef]
- Košinová, P.; Berka, K.; Wykes, M.; Otyepka, M.; Trouillas, P. Positioning of Antioxidant Quercetin and Its Metabolites in Lipid Bilayer Membranes: Implication for Their Lipid-Peroxidation Inhibition. J. Phys. Chem. B 2012, 116, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Raghunathan, M.; Zubovski, Y.; Venable, R.M.; Pastor, R.W.; Nagle, J.F.; Tristram-Nagle, S. Structure and elasticity of lipid membranes with genistein and daidzein bioflavonoids using X-ray scattering and MD simulations. J. Phys. Chem. B 2012, 116, 3918–3927. [Google Scholar] [CrossRef] [PubMed]
- Dwiecki, K.; Neunert, G.; Polewski, P.; Polewski, K. Antioxidant activity of daidzein, a natural antioxidant, and its spectroscopic proterties in organic solvents and phosphatidylcholine liposomes. J. Photochem. Photobiol. B 2009, 96, 242–248. [Google Scholar] [CrossRef]
- Pawlikowska-Pawlęga, B.; Dziubińska, H.; Król, E.; Trębacz, K.; Jarosz-Wilkołazka, A.; Paduch, R.; Gawron, A.; Gruszecki, W.I. Characteristics of quercetin interactions with liposomal and vacuolar membranes. Biochim. Biophys. Acta-Biomembr. 2014, 1838, 254–265. [Google Scholar] [CrossRef]
- Pawlikowska-Pawlęga, B.; Misiak, L.E.; Zarzyka, B.; Paduch, R.; Gawron, A.; Gruszecki, W.I. Localization and interaction of genistein with model membranes formed with dipalmitoylphosphatidylcholine (DPPC). Biochim. Biophys. Acta Biomembr. 2012, 1818, 1785–1793. [Google Scholar] [CrossRef]
- Bărbînţă Pătraşcu, M.E.; Drăguşin, M.; Ţugulea, L.; Meghea, A. Effect of quercetin on artificial lipid membranes. UPB Sci. Bull. 2009, 71, 41–50. [Google Scholar]
- Wójtowicz, K.; Pawlikowska-Pawlęga, B.; Gawron, A.; Gruszecki, W.I. Modifying effect of quercetin on the lipid membrane. Folia Histochem. Cytobiol. 1996, 34, 49–50. [Google Scholar]
- Wesołowska, O.; Hendrich, A.B.; Lania-Pietrzak, B.; Wisniewski, J.; Molnar, J.; Ocsovszki, I.; Michalak, K. Perturbation of the lipid phase of a membrane is not involved in the modulation of MRP1 transport activity by flavonoids. Cell. Mol. Biol. Lett. 2009, 14, 199–221. [Google Scholar] [CrossRef]
- Selvaraj, S.; Krishnaswamy, S.; Devashya, V.; Sethuraman, S. Influence of membrane lipid composition on flavonoid-membrane interactions: Implications on their biological activity. Prog. Lipid Res. 2015, 58, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, H. Membrane interactions of phytochemicals as their molecular mechanism applicable to the discovery of drug leads from plants. Molecules 2015, 20, 18923–18966. [Google Scholar] [CrossRef] [PubMed]
- Eid, J.; Jraij, A.; Greige-Gerges, H.; Monticelli, L. Effect of quercetin on lipid membrane rigidity: Assessment by atomic force microscopy and molecular dynamics simulations. Biochim. Biophys. Acta Adv. 2021, 1, 100018. [Google Scholar] [CrossRef] [PubMed]
- de Granada-Flor, A.; Sousa, C.; Filipe, H.A.L.; Santos, M.S.C.S.; de Almeida, R.F.M. Quercetin dual interaction at the membranę level. Chem. Commun. 2019, 55, 1750–1753. [Google Scholar] [CrossRef]
- Strawa, J.W.; Jakimiuk, K.; Szoka, Ł.; Brzezinski, K.; Drozdzal, P.; Pałka, J.A.; Tomczyk, M. New polymethoxyflavones from Hottonia palustris evoke DNA biosynthesis-inhibitory activity in an oral squamous carcinoma (SCC-25) cell line. Molecules 2022, 27, 4415. [Google Scholar] [CrossRef]
- Jakimiuk, K.; Strawa, J.W.; Granica, S.; Tomczyk, M. New flavone C-glycosides from Scleranthus perennis and their anti-collagenase activity. Molecules 2021, 26, 5631. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Name | MW (g/mol) | LogP | pKa Acidic | LogD (pH 4) |
|---|---|---|---|---|---|
| HZ4 | 5-hydroxy-2′,6′-dimethoxyflavone (C17H14O5) | 298.294 | 2.18 | 6.29 | 1.9 |
| SP9 | 8-C-β-D-xyloside-(2′′-O-(4′′′-acetoxy)glucoso)-5,7-dihydroxy-3′-methoxy-4′-acetoksyflawone (C31H34O17) | 678.596 | −2.42 | 5.02 | −3.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapral-Piotrowska, J.; Strawa, J.W.; Jakimiuk, K.; Wiater, A.; Tomczyk, M.; Gruszecki, W.I.; Pawlikowska-Pawlęga, B. Investigation of the Membrane Localization and Interaction of Selected Flavonoids by NMR and FTIR Spectroscopy. Int. J. Mol. Sci. 2023, 24, 15275. https://doi.org/10.3390/ijms242015275
Kapral-Piotrowska J, Strawa JW, Jakimiuk K, Wiater A, Tomczyk M, Gruszecki WI, Pawlikowska-Pawlęga B. Investigation of the Membrane Localization and Interaction of Selected Flavonoids by NMR and FTIR Spectroscopy. International Journal of Molecular Sciences. 2023; 24(20):15275. https://doi.org/10.3390/ijms242015275
Chicago/Turabian StyleKapral-Piotrowska, Justyna, Jakub W. Strawa, Katarzyna Jakimiuk, Adrian Wiater, Michał Tomczyk, Wiesław I. Gruszecki, and Bożena Pawlikowska-Pawlęga. 2023. "Investigation of the Membrane Localization and Interaction of Selected Flavonoids by NMR and FTIR Spectroscopy" International Journal of Molecular Sciences 24, no. 20: 15275. https://doi.org/10.3390/ijms242015275