
Myxozoan Ceratomyxids Infecting the Gallbladder of Amazonian Ornamental Cichlid Fish: Description of Ellipsomyxa santarenensis n. sp. and Report of Ceratomyxa amazonensis in a New Host


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Collections and Parasite Sampling
2.2. Morphological Analyses
2.3. DNA-SSU rDNA Sequencing
2.4. Phylogenetic Analyses
3. Results
3.1. Myxozoa Parasite of Geophagus altifrons Heckel, 1840
3.1.1. Morphological Data
3.1.2. SSU rDNA Data
3.2. Myxozoa Parasite of Satanoperca jurupari (Heckel, 1840)
3.2.1. Morphological Data
3.2.2. SSU rDNA Data
3.3. Phylogenetic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Okamura, B.; Hartigan, A.; Naldoni, J. Extensive uncharted biodiversity: The parasite dimension. Integr. Comp. Biol. 2018, 58, 1132–1145. [Google Scholar] [CrossRef] [PubMed]
- Okamura, B.; Gruhl, A.; Bartholomew, J.L. An Introduction to Myxozoan Evolution, Ecology and Development; Springer International Publisher: Cham, Switzerland, 2015; pp. 69–84. [Google Scholar]
- Eiras, J.C. Synopsis of the species of the genus Ceratomyxa Thelohan, 1892 (Myxozoa: Myxosporea: Ceratomyxidae). Syst. Parasitol. 2006, 65, 49–71. [Google Scholar] [CrossRef] [PubMed]
- Eiras, J.C.; Cruz, C.; Saraiva, A. Synopsis of the species of Ceratomyxa Thelohan, 1892 (Cnidaria, Myxosporea, Ceratomyxidae) described between 2007 and 2017. Syst. Parasitol. 2018, 95, 427–446. [Google Scholar] [CrossRef]
- Azevedo, C.; Rocha, S.; Casal, G.; São Clemente, C.S.; Matos, P.S.; Al-Quraishy, A.; Matos, E. Light and ultrastructural description of Meglitschia mylei n. sp. (Myxozoa) from Myleus rubripinnis (Teleostei: Serrasalmidae) in the Amazon River System. J. Eukaryot. Microbiol. 2011, 58, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, C.; Rocha, S.; Casal, G.; São Clemente, C.S.; Matos, P.S.; Al-Quraishy, A.; Matos, E. Ultrastructural description of Ceratomyxa microlepis sp. nov. (Phylum Myxozoa): A parasite infecting the gall bladder of Hemiodus microlepis, a freshwater teleost from the Amazon River. Mem. Inst. Oswaldo Cruz. 2013, 108, 150–154. [Google Scholar] [CrossRef] [Green Version]
- Mathews, P.D.; Naldoni, J.; Maia, A.A.M.; Adriano, E.A. Morphology and small subunit rDNA-based phylogeny of Ceratomyxa amazonensis n. sp. parasite of Symphysodon discus, an ornamental freshwater fish from Amazon. Parasitol. Res. 2016, 115, 4021–4025. [Google Scholar] [CrossRef]
- Adriano, E.A.; Okamura, B. Motility, morphology and phylogeny of the plasmodial worm, Ceratomyxa vermiformis n. sp. (Cnidaria: Myxozoa: Myxosporea). Parasitology 2017, 144, 158–168. [Google Scholar] [CrossRef] [Green Version]
- Zatti, S.A.; Atkinson, S.D.; Bartholomew, J.L.; Maia, A.A.M.; Adriano, E.A. Ceratomyxa gracillima n. sp. (Cnidaria: Myxosporea) provides evidence of panmixia and ceratomyxid radiation in the Amazon basin. Parasitology 2018, 145, 1137–1146. [Google Scholar] [CrossRef]
- Silva, M.S.; Carvalho, A.E.F.B.; Hamoy, I.; Matos, E.R. Coelozoic parasite of the family Ceratomyxidae (Myxozoa, Bivalvulida) described from motile vermiform plasmodia found in Hemiodus unimaculatus Bloch, 1794. Parasitol. Res. 2020, 119, 871–878. [Google Scholar] [CrossRef]
- Bittencourt, L.S.; Silva, D.T.; Hamoy, I.; Carvalho, A.A.; Silva, M.F.; Videira, M.; Carvalho, J.C.T.; Matos, E.R. Morphological and Phylogenetic Features of Ceratomyxa macapaensis n. sp. (Myxozoa: Ceratomyxidae) in Mesonauta festivus Heckel, 1840 (Cichliformes: Cichlidae) from the eastern Amazon region. Acta Parasitol. 2021, 67, 322–329. [Google Scholar] [CrossRef]
- Zatti, S.A.; Adriano, E.A.; Araújo, B.L.; Franzolin, G.N.; Maia, A.A.M. Expanding the geographic distribution of the freshwater parasite Ceratomyxa (Cnidaria: Myxozoa) with vermiform-type plasmodia. Microb. Pathog. 2022, 162, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Araújo, B.L.; Adriano, E.A.; Franzolin, G.N.; Zatti, S.A.; Naldoni, J. A novel Ceratomyxa species (Myxozoa: Cnidaria) infecting an Amazonian catfish. Parasitology 2022, 89, 102582. [Google Scholar] [CrossRef] [PubMed]
- Franzolin, G.N.; Araújo, B.L.; Zatti, S.A.; Naldoni, J.; Adriano, E.A. Occurrence of the host-parasite system Rhaphiodon vulpinus and Ceratomyxa barbata n. sp. in the two largest watersheds in South America. Parasitol. Int. 2022, 91, 102651. [Google Scholar] [CrossRef]
- Adriano, E.A.; Zatti, S.A.; Okamura, B. How to build single-celled cnidarians with worm-like motility: Lessons from Myxozoa. J. Anat. 2022, 240, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Zatti, S.A.; Atkinson, S.D.; Maia, A.A.; Corrêa, L.L.; Bartholomew, J.L.; Adriano, E.A. Novel Myxobolus and Ellipsomyxa species (Cnidaria: Myxozoa) parasiting Brachyplatystoma rousseauxii (Siluriformes: Pimelodidae) in the Amazon basin, Brazil. Parasitol. Int. 2018, 67, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Zatti, S.A.; Maia, A.A.; Adriano, E.A. Growing diversity supports radiation of an Ellipsomyxa lineage into the Amazon freshwater: Description of two novel species parasitizing fish from Tapajós and Amazon rivers. Acta Trop. 2020, 211, 105616. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.T.; Matos, P.S.; Lima, A.M.; Furtado, A.P.; Hamoy, I.; Matos, E.R. Ellipsomyxa arariensis n. sp. (Myxozoa: Ceratomyxidae), a new myxozoan parasite of Pygocentrus nattereri Kner, 1858 (Teleostei: Characidae) and Pimelodus ornatus Kner, 1858 (Teleostei: Pimelodidae) from Marajó Island, in the Brazilian Amazon region. Parasitol. Res. 2018, 117, 3537–3545. [Google Scholar] [CrossRef]
- Ferreira, R.L.S.; Silva, D.T.; Carvalho, A.A.; Bittencourt, L.S.; Hamoy, I.; Matos, E.; Videira, M. Ellipsomyxa tucujuensis n. sp. (Myxozoa: Ceratomyxidae), parasita de Satanoperca jurupari (Osteichthyes: Cichlidae) da Amazônia brasileira. Parasitol. Int. 2021, 83, 102332. [Google Scholar] [CrossRef]
- Silvano, R.A.; Nitschke, P.P.; Vieira, K.C.; Nagl, P.; Martínez, A.T.; Dutra, M.C.; Andrade, M.C. Atlas of Fish of Tapajós and Negro Rivers III: Perciformes and Other Fish Groups. In Fish and Fisheries in the Brazilian Amazon; Springer: Cham, Switzerland, 2020; pp. 321–414. [Google Scholar]
- Froese, R.; Pauly, D. World Wide Web Electronic Publication. FishBase. 2022. Available online: www.fishbase.org (accessed on 4 April 2023).
- Lom, J.; Arthur, J. A guideline for the preparation of species descriptions in Myxosporea. J. Fish Dis. 1989, 12, 151–156. [Google Scholar] [CrossRef]
- Gunter, N.L.; Whipps, C.N.; Adlard, R.D. Ceratomyxa (Myxozoa: Bivalvulida): Robust taxon or genus of convenience. Int. J. Parasitol. 2009, 39, 1395–1405. [Google Scholar] [CrossRef]
- Whipps, C.M.; Adlard, R.D.; Bryant, M.S.; Lester, R.J.G.; Findlay, V.; Kent, M.L. First report of three Kudoa species from Eastern Australia: Kudoa thyrsites from Mahi mahi (Coryphaena hippurus), Kudoa amamiensis and Kudoa minithyrsites n. sp. from Sweeper (Pempheris ypsilychnus). J. Eukaryot. Microbiol. 2003, 50, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Heiniger, H.; Adlard, R.D. Relatedness of novel species of Myxidium Bütschli, 1882, Zschokkella Auerbach, 1910 and Ellipsomyxa Køie, 2003 (Myxosporea: Bivalvulida) from the gall bladders of marine fishes (Teleostei) from Australian waters. Syst. Parasitol. 2014, 87, 47–72. [Google Scholar] [CrossRef] [PubMed]
- Thabet, A.; Mansour, L.; Al Omar, S.Y.; Tlig-Zouari, S. Ceratomyxa tunisiensis n. sp. (Myxosporea: Bivalvulida) from the gallbladders of two carangid fish caught off the coast of Tunisia. J. Eukaryot. Microbiol. 2016, 63, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q. Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef] [PubMed]
- Barta, J.R.; Martin, D.S.; Liberator, P.A.; Dashkevicz, M.; Anderson, J.W.; Feighner, S.D.; Elbrecht, A.; Perkins-Barrow, A.; Jenkins, M.C.; Danforth, H.D.; et al. Phylogenetic relationships among eight Eimeria species infecting domestic fowl inferred using complete small subunit ribosomal DNA sequences. J. Parasitol. 1997, 83, 262–271. [Google Scholar] [CrossRef]
- Molnár, K.; Eszterbauer, E.; Székely, C.; Dan, A.; Harrach, B. Morphological and molecular biological studies on intramuscular Myxobolus spp. of cyprinid fish. J. Fish Dis. 2002, 25, 643–652. [Google Scholar] [CrossRef]
- Edgar, R.C. Muscle: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Drummond, A. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Xia, X. DAMBE5: A comprehensive software package for data analysis in molecular biology and evolution. Mol. Biol. Evol. 2013, 30, 1720–1728. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, K. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: Piscataway, NJ, USA, 2010; pp. 1–8. [Google Scholar]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. Molecular Evolution, Phylogenetics and Epidemiology: Fig-Tree. 2009. Available online: http.ac.uk/software/figtree/ (accessed on 10 November 2015).
- Zatti, S.A.; Atkinson, S.D.; Bartholomew, J.L.; Maia, A.A.M.; Adriano, E.A. Amazonian waters harbour an ancient freshwater Ceratomyxa lineage (Cnidaria: Myxosporea). Acta Trop. 2017, 169, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Sousa, F.B.; Milanin, T.; Morandini, A.C.; Espinoza, L.L.; Flores-Gonzales, A.; Gomes, A.L.S.; Mathews Delgado, P. Molecular diagnostic based on 18S rDNA and supplemental taxonomic data of the cnidarian coelozoic Ceratomyxa (Cnidaria, Myxosporea) and comments on the intraspecific morphological variation. Zoosystematics Evol. 2021, 97, 307–314. [Google Scholar] [CrossRef]
- Zhai, Y.; Whipps, C.M.; Gu, Z.; Guo, Q.; Wu, Z.; Wang, H.; Liu, Y. Intraspecific morphometric variation in myxosporeans. Folia Parasitol. 2016, 63, 001. [Google Scholar] [CrossRef] [Green Version]
- Mathews, P.D.; Mertins, O.; Pereira, J.O.L.; Maia, A.A.M.; Adriano, E.A. Morphology and 18S rDNA sequencing of Henneguya peruviensis n. sp. (Cnidaria: Myxosporea), a parasite of the Amazonian ornamental fish Hyphessobrycon loretoensis from Peru: A myxosporean dispersal approach. Acta Tropica. 2018, 187, 207–213. [Google Scholar] [CrossRef]
- Biosecurity, N.Z. Import Risk Analysis: Ornamental Fish; Ministry of Agriculture and Forestry: Wellington, New Zealand, 2005. [Google Scholar]
- Hallett, S.L.; Hartigan, A.; Atkinson, S.D. Myxozoans on the move: Dispersal modes, exotic species and emerging diseases. In Myxozoan Evolution, Ecology and Development; Springer International Publisher: Cham, Switzerland, 2015; pp. 343–362. [Google Scholar]
- Figueredo, R.T.A.; Müller, M.I.; Arana, S.; Long, P.F.; Adriano, E.A. Phylogenetic and host-parasite relationship analyses of Henneguya caquetaia sp. nov (Myxosporea: Myxobolidae) infecting an Amazonian cichlid fish. Microb. Pathog. 2023, 179, 106116. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
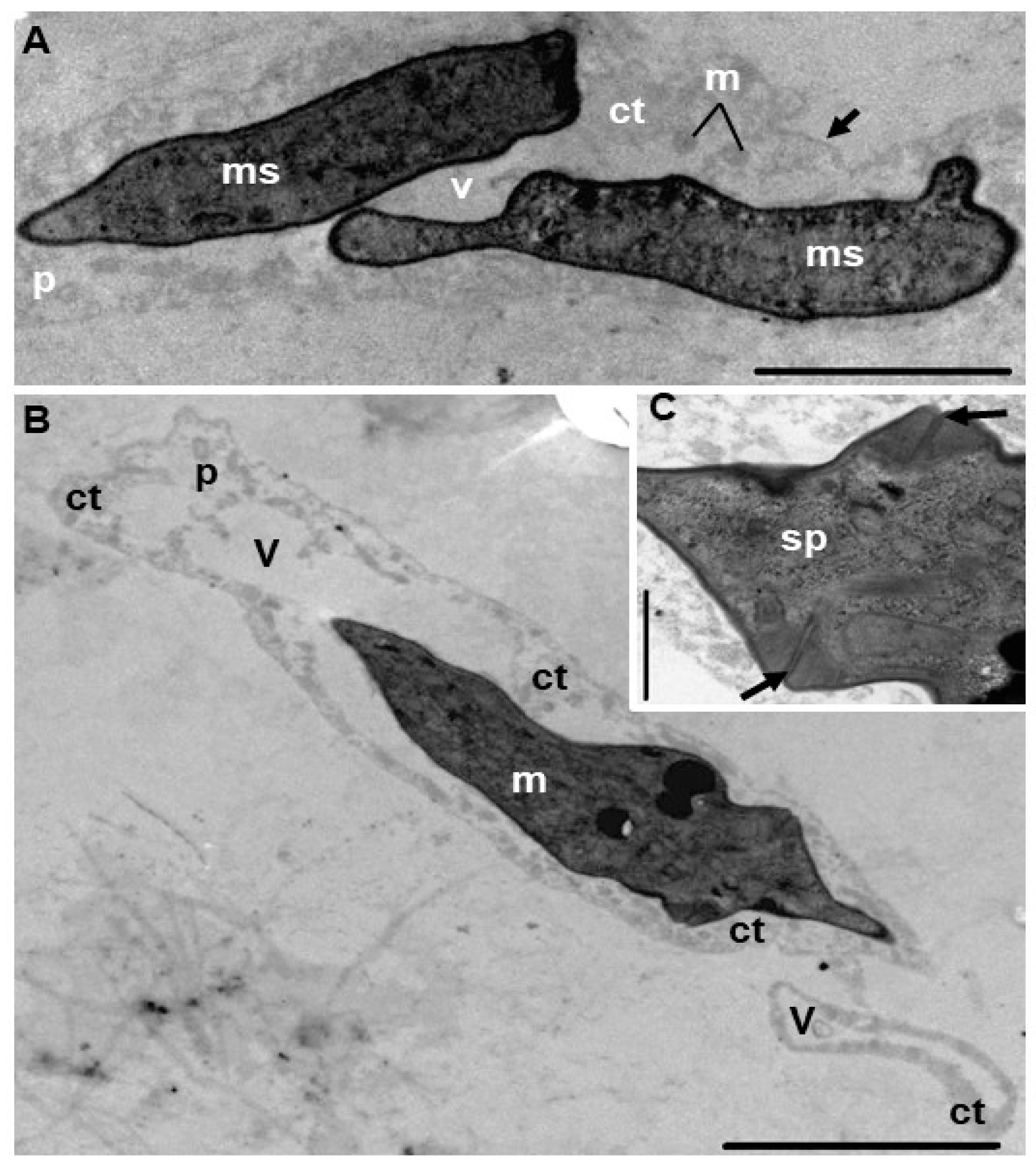{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Ceratomyxa mylei (syn. Meglitschia mylei) * | Ceratomyxa microlepis | Ceratomyxa amazonensis | Ceratomyxa vermiformis | Ceratomyxa brasiliensis | Ceratomyxa gracillima | Ceratomyxa fonsecai | Ceratomyxa macapaensis | Ceratomyxa cf. fonsecai | Ceratomyxa mandii | Ceratomyxa barbata | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Characters | ||||||||||||||
| Plasmodia shape | -- | -- | -- | Worm-like | Worm-like | Worm-like | Worm-like | Worm-like | Worm-like | Worm-like | Worm-like | Worm-like | Worm-like | |
| Immature stages | ||||||||||||||
| Length | -- | -- | -- | -- | 71.6 (22.1–188.1) | 112 (72–134) | -- | -- | 494.2 (432.5–542.2) | 112.6 (82–141) | 184.4 (96–204) | |||
| Width | -- | -- | -- | -- | 8.9 (5.7–20.1) | 8.9 (4·6–11.8) | -- | -- | 28.1 (21.8–34.4) | 5.1 (4.2–6.5) | 8.17 (5.32–12.39) | |||
| Mature stages | ||||||||||||||
| Length | -- | -- | -- | 62.3 (58.4–64.2) | 106.9 (67.3–202.6) | 442 (379–520) | 304.2 (196–402) | 181 (173–246) | 484 (445–523) | -- | 484.7 (402–556) | 199.3 (174–221) | 288 (235.5–343.1) | |
| Width | -- | -- | -- | 7.8 (6.6–8.9) | 7.6 ± 1.7 (4.9–11.1) | 22.1 (18–26) | 35.7 (18.3–55.1) | 11.4 (7.2–16.2 | -- | -- | 37.8 (28.6–52.5) | 10.5 (7.9–12.9) | 18 (12.3–25.9) | |
| Myxospores | ||||||||||||||
| Shape | strongly arcuate | elongated/lightly arcuate | arcuate | elongated/lightly arcuate | elongated/lightly arcuate | strongly arcuate | elongated/lightly arcuate | strongly arcuate | elongated/lightly arcuate | elongated/lightly arcuate | elongated/lightly arcuate | elongated/lightly arcuate | elongated/lightly arcuate | |
| Length | 5 | 5.2 ± 0.4 | 7.0 (6.2–7.6) | 4.72 (4.52–4.81) | 4.9 ± 1.4(3.1–6.3) | 4.5 (4.8–4.2) | 6.3 (5.1–7.5) | 4.4 (3.0–5.7) | 2.6 (2.3–2.8) | 4.2 ± 0.5 | 3.3 (2.9–3.9) | 4.6 (3.4–5.5) | 2.9 (2.1–3.8) | |
| Thickness | 9.1 | 35.5 ± 0.9 | 15.8 (15.0–16.7) | 24.2 (23.9–25.3) | 23.9 (18.8–28.6) | 8.4 (7.9–9.3) | 41.2 (37.1–47.6) | 7.0 (6.0–8.2). | 28.9 (24.3–32.5) | 22.75 ± 0.3 | 28 (24.7–31.7) | 31.2 (26.2–36.3) | 21.7 (17.6–29.9) | |
| Posterior angle | 30° | 58°–60°(?) | 105–115° | 154° (153°–156°) | 147.8° (174.9°–144.7°) | 30.2° (22–43) | 147° | 35°–40° | 164.8° | -- | 166° (146°–179°) | 162° (143–178) | 164° (139–178) | |
| Nematocysts | ||||||||||||||
| Length | 2.1 ± 0.3 | 2.2 ± 0.3 | 3.2 (2.4–3.6) | 2.31 (2.29–2.33) | 2.4 (1.2–5.6) | 2.7 (2.5–2.9) | 2.6 (2–3.3) | 1.9 (1.5–2.5) | 1.9 (1.5–2.3) | 1.63 ± 0.1 | 1.59 (1.11–2.35) | 1.8 (1.0–2.5) | 1.43 (1.1–1.5) | |
| Width | 2.1 ± 0.3 | 2.2 ± 0.3 | 2.6 (2.4–2.9) | 2.1 (2.13–2.17) | 1.9 (1.5–2.8) | 2.7 (2.5–2.9) | 2.5 (1.8–3.7) | 1.9 (1.5–2.5) | 1.7 (1.4–2.1) | 1.86 ± 0.3 | 1.47 (0.9–2.11) | 1.9 (1.2–2.4) | 1.6 (1.1–2.3) | |
| Tubule turns | 5–6 | 5–6 | 3–4 | 3–4 | 3–4 | 3–4 | 3–4 | 2–3 | 3–4 | 3–4 | -- | 3–4 | 3 | |
| Host | Myloplus rubripinnis (Müller & Troschel, 1844) | Hemiodus microlepis Kner, 1858 | Symphysodon discus Heckel, 1840 | Symphysodon discus Heckel, 1840 | Geophagus altifrons Heckel, 1840 | Colossoma macropomum Cuvier, 1816 | Cichla monoculus Agassiz, 1831 | Brachyplatystoma rousseauxii (Castelnau, 1855) | Hemiodus unimaculatus Bloch, 1794 | Mesonauta festivus Heckel, 1840 | Hemiodus orthonops Eigenmann and Kennedy, 1903 | Pimelodina flavipinnis Steindachner, 1876 | Rhaphiodon vulpinus Spix & Agassiz, 1829 | |
| Referências | [5] | [6] | [7] | 39 | This study | [8] | [38] | [9] | [10] | [11] | [12] | [13] | [14] | |
| Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1-Ceratomyxa amazonensis (this study) | ||||||||||
| 2-Ceratomyxa amazonensis KX236169 | 0.6 | |||||||||
| 3-Ceratomyxa amazonensis MN064752 | 0.2 | 0.1 | ||||||||
| 4-Ceratomyxa mandi MZ504285 | 8.3 | 9.0 | 8.7 | |||||||
| 5-Ceratomyxa barbata MZ504286 | 5.5 | 5.5 | 5.9 | 7.0 | ||||||
| 6-Ceratomyxa brasiliensis KU978813 | 2.5 | 2.5 | 3.0 | 8.7 | 5.9 | |||||
| 7-Ceratomyxa gracillima KY934184 | 7.8 | 8.4 | 8.0 | 4.6 | 7.2 | 7.8 | ||||
| 8-Ceratomyxa vermiformis KX278420 | 6.1 | 6.0 | 5.1 | 6.8 | 3.7 | 6.2 | 6.9 | |||
| 9-Ceratomyxa macapaenses MT939250 | 5.3 | 5.4 | 5.5 | 7.0 | 4.7 | 6.1 | 7.9 | 0.7 | ||
| 10-Ceratomyxa cf fonsecai MW053456 | 7.7 | 7.6 | 8.0 | 6.8 | 5.9 | 7.8 | 6.4 | 6.3 | 6.5 | - |
| Species | Myxospore | Nematocyst | Host and Locality | |||
|---|---|---|---|---|---|---|
| Length | Width | Length | Width | Tubule Turns | ||
| Ellipsomyxa santarenensis n. sp. [this study] | 12.0 (10.7–13.7) | 7.6 (5.6–8.2) | 2.8 (2.0–3.6) | 2.8 (2.1–3.7) | 4–5 | Satanoperca jurupari Amazon and Tapajós River, PA |
| Ellipsomyxa tucujuensis [19] | 10.11 ± 0.86 | 7.81 ± 1.14 | 3.12 ± 0.53 | 2.5 ± 0.32 | 5–6 | Satanoperca jurupari Curiau River, AP |
| Ellipsomyxa paraensis [17] | 11.5 (10.5–12.4) | 7.5 (6.6–8.6) | 3.2 (2.1–3.9) | 2.6 (2–3.3) | 2–3 | Cichla monoculus Amazon River, PA |
| Ellipsomyxa plagioscioni [17] | 11.1 (10.2–12.8) | 6.6 (5.6–8.6) | 3.8 (3.2–4.4) | 2.8 (2.3–3.3) | 5–6 | Plagioscion squamosissimus Tapajós River, PA |
| Ellipsomyxa amazonensis [16] | 12.8 (12.3–13.6) | 7.6 (6.7–8.7) | 3.8 (3.8–4.0) | 3.1 (2.5–3.4) | 2–3 | Brachyplatystoma rousseauxii Tapajós River, PA |
| Ellipsomyxa arariensis [18] | 12.6 (12.3–13.6) | 7.3 (6.7–8.7) | 3.5 (3.8–4.0) | 2.6 (2.5–3.4) | 5–6 | Pigocentrus nattereri Arari River, PA |
| Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| 1-Ellipsomyxa santarenensis n. sp. (this study) | |||||||
| 2-Ellipsomyxa amazonensis MF193889 | 1.6 | ||||||
| 3-Ellipsomyxa plagioscioni MT039013 | 6.3 | 6.5 | |||||
| 4-Ellipsomyxa paraenses MH364399 | 1.6 | 0.3 | 5.2 | ||||
| 5-Ellipsomyxa arariensis MH30806 | 2.5 | 2.7 | 7.4 | 2.5 | |||
| 6-Ellipsomyxa arariensis MH183026 | 1.9 | 2.9 | 6.8 | 1.9 | 0.5 | ||
| 7-Ellipsomyxa tucujuensis MN999871 | 8.8 | 9.2 | 12.6 | 7.4 | 10.5 | 11.5 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueredo, R.T.A.; Müller, M.I.; Long, P.F.; Adriano, E.A. Myxozoan Ceratomyxids Infecting the Gallbladder of Amazonian Ornamental Cichlid Fish: Description of Ellipsomyxa santarenensis n. sp. and Report of Ceratomyxa amazonensis in a New Host. Diversity 2023, 15, 830. https://doi.org/10.3390/d15070830
Figueredo RTA, Müller MI, Long PF, Adriano EA. Myxozoan Ceratomyxids Infecting the Gallbladder of Amazonian Ornamental Cichlid Fish: Description of Ellipsomyxa santarenensis n. sp. and Report of Ceratomyxa amazonensis in a New Host. Diversity. 2023; 15(7):830. https://doi.org/10.3390/d15070830
Chicago/Turabian StyleFigueredo, Rayline T. A., Maria I. Müller, Paul F. Long, and Edson A. Adriano. 2023. "Myxozoan Ceratomyxids Infecting the Gallbladder of Amazonian Ornamental Cichlid Fish: Description of Ellipsomyxa santarenensis n. sp. and Report of Ceratomyxa amazonensis in a New Host" Diversity 15, no. 7: 830. https://doi.org/10.3390/d15070830