
Predicting the Potential Geographic Distribution of Invasive Freshwater Apple Snail Pomacea canaliculate (Lamarck, 1819) under Climate Change Based on Biomod2

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Occurrence Records of P. canaliculata
2.2. Environmental Variables Related to P. canaliculata
2.3. Construction and Evaluation of SDMs
2.4. Division of the PGDs and Calculation of Centroids
3. Results
3.1. Evaluation of SDMs
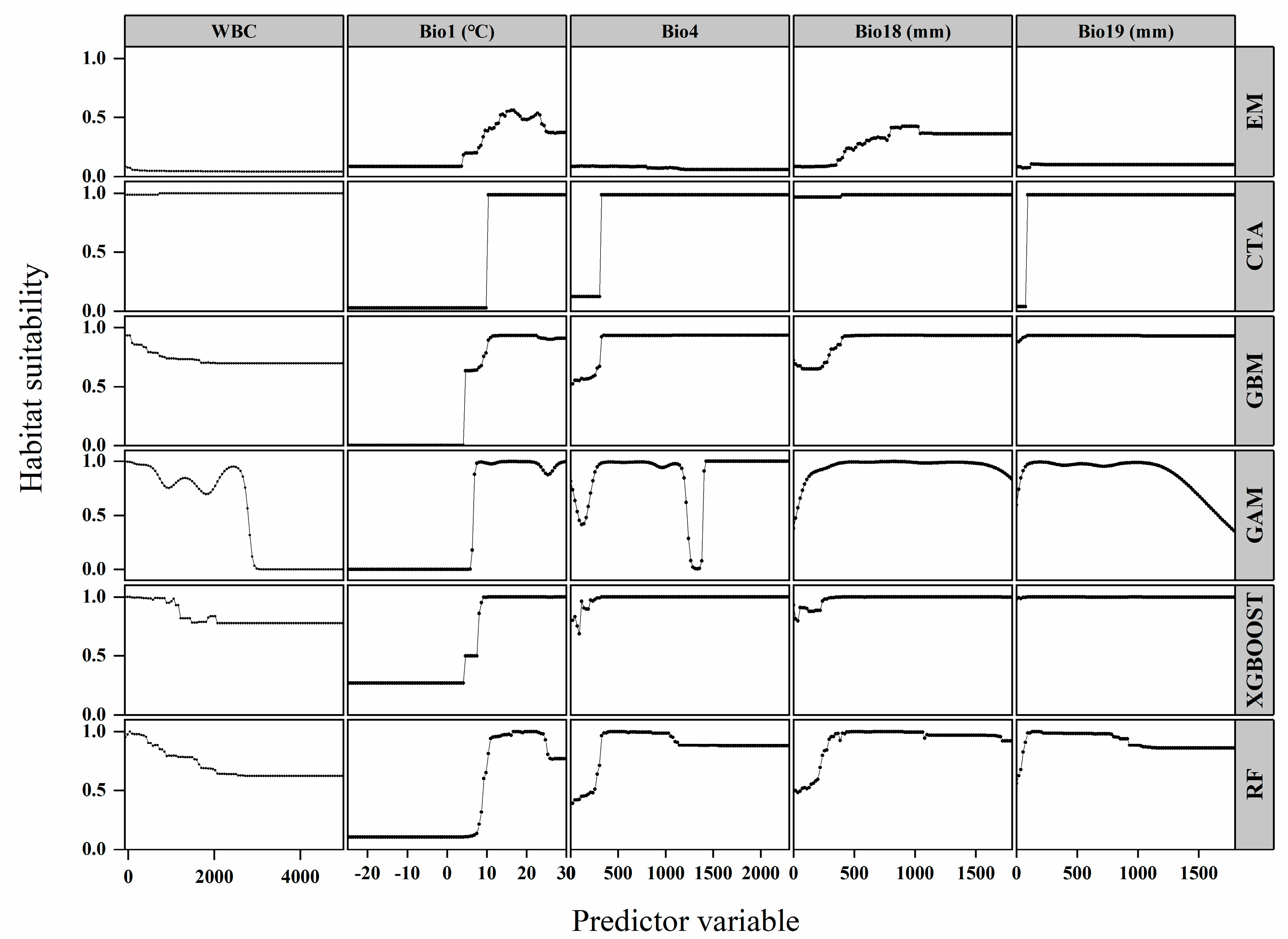
3.2. Evaluation of Environmental Variables
3.3. Occurrence Records and PGDs of P. canaliculata in China under the Current Climate Conditions
3.4. PGDs of P. canaliculata under Future Climate Conditions
3.5. Centroids Shifts of P. canaliculata under Future Climate Change Conditions
4. Discussion
4.1. Model Evaluation
4.2. PGDs of P. canaliculata
4.3. Environmental Variables
4.4. PGDs under Climate Change
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wan, F.; Guo, J.; Wang, D. Alien invasive species in China: Their damages and management strategies. Biodivers. Sci. 2002, 10, 119–125. [Google Scholar]
- Biondi, A.; Guedes, R.N.C.; Wan, F.H.; Desneux, N. Ecology, worldwide spread, and management of the invasive South American tomato pinworm, Tuta absoluta: Past, present, and future. Annu. Rev. Entomol. 2018, 63, 239–258. [Google Scholar] [CrossRef]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Gonçalves da Silva, A.; Kolokotronis, S.O.; Wharton, D. Modeling the eradication of invasive mammals using the sterile male technique. Biol. Invasions 2010, 12, 751–759. [Google Scholar] [CrossRef]
- Simberloff, D. How much information on population biology is needed to manage introduced species? Conserv. Biol. 2003, 17, 83–92. [Google Scholar] [CrossRef]
- Liu, X.; Blackburn, T.M.; Song, T.; Li, X.; Huang, C.; Li, Y. Risks of biological invasion on the belt and road. Curr. Biol. 2019, 29, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.H.; Yang, N.W. Invasion and management of agricultural alien insects in China. Annu. Rev. Entomol. 2016, 61, 77–98. [Google Scholar] [CrossRef]
- Luque, G.M.; Bellard, C.; Bertelsmeier, C.; Bonnaud, E.; Genovesi, P.; Simberloff, D.; Courchamp, F. The 100th of the world’s worst invasive alien species. Biol. Invasions 2014, 16, 981–985. [Google Scholar] [CrossRef]
- Mochida, O. Spread of freshwater Pomacea snails (Pilidae, Mollusca) from Argentina to Asia. Micronesica 1991, 3, 51–62. [Google Scholar]
- Naylor, R. Invasions in agriculture: Assessing the cost of the golden apple snail in Asia. Ambio 1996, 25, 443–448. [Google Scholar]
- Adalla, C.; Magsino, E. Understanding the golden apple snail (Pomacea canaliculata): Biology and early initiatives to control the pest in the Philippines. In Global Advances in Ecology and Management of Golden Apple Snails; Philippine Rice Research Institute (PhilRice): Los Baños, Philippines, 2006; pp. 199–213. [Google Scholar]
- Sawangproh, W.; Poonswad, P. Population dynamics of Pomacea canaliculata (Lamarck, 1822) in relation to rice cultivation practice and seasons in Nakhon Pathom, Thailand. KKU Sci. J. 2010, 38, 228–238. [Google Scholar]
- Yahaya, H.; Nordin, M.; Muhamad, H.; Sivapragasam, A. Golden apple snails in Malaysia. In Global Advances in Ecology and Management of Golden Apple Snails; Philippine Rice Research Institute (PhilRice): Los Baños, Philippines, 2006; pp. 215–230. [Google Scholar]
- Duong, N.C. The golden apple snail in Vietnam. In Global Advances in Ecology and Management of Golden Apple Snails; Philippine Rice Research Institute (PhilRice): Los Baños, Philippines, 2006; pp. 243–254. [Google Scholar]
- Carlsson, N.O.; Lacoursiere, J.O. Herbivory on aquatic vascular plants by the introduced golden apple snail (Pomacea canaliculata) in Lao PDR. Biol. Invasions 2005, 7, 233–241. [Google Scholar] [CrossRef]
- Carlsson, N.O.; Brönmark, C. Size-dependent effects of an invasive herbivorous snail (Pomacea canaliculata) on macrophytes and periphyton in Asian wetlands. Freshw. Biol. 2006, 51, 695–704. [Google Scholar] [CrossRef]
- Fang, L.; Wong, P.K.; Lin, L.; Lan, C.; Qiu, J. Impact of invasive apple snails in Hong Kong on wetland macrophytes, nutrients, phytoplankton and filamentous algae. Freshw. Biol. 2010, 55, 1191–1204. [Google Scholar] [CrossRef]
- Cowie, R.H. Angiostrongylus cantonensis: Agent of a sometimes fatal globally emerging infectious disease (rat lungworm disease). ACS Chem. Neurosci. 2017, 8, 2102–2104. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, N.S.; Bhadelia, N. Infections associated with exotic cuisine: The dangers of delicacies. Microbiol. Spectr. 2015, 3, 355–374. [Google Scholar] [CrossRef] [PubMed]
- Cowie, R.H. Apple snails (Ampullariidae) as agricultural pests: Their biology, impacts and management. In Molluscs as Crop Pests; CABI Publishing: Wallingford, UK, 2002; p. 145192. [Google Scholar]
- Marwoto, R.M.; Isnaningsih, N.R.; Joshi, R.C. The invasive apple snail (Pomacea spp.) in Indonesia. Agric. Dev. 2018, 35, 41–48. [Google Scholar]
- Salleh, N.H.M.; Arbain, D.; Daud, M.Z.M.; Pilus, N.; Nawi, R. Distribution and management of Pomacea canaliculata in the northern region of Malaysia: Mini review. APCBEE Procedia 2012, 2, 129–134. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, Y.; Li, X.; Wang, X.; Mu, X.; Song, H.; Wang, P.; Liu, C.; Liu, C.; Luo, J. Historical invasion, expansion process and harm investigation of Pomacea canaliculata in China. Sci. Agric. Sic. Bull. 2010, 26, 245–250. [Google Scholar]
- Baker, R.; Candresse, T.; sné Simon, E.; Gilioli, G.; Grégoire, J.; Jeger, M.; Karadjova, O.; Lövei, G.; Makowski, D.; Manceau, C. Statement on the identity of apple snails. EFSA J. 2012, 10, 1–11. [Google Scholar]
- Lucero, J.M. Regional Expansion and Evaluation of Potential Chemical Control for Invasive Apple Snails (Pomacea maculata) in Southwest Louisiana; Louisiana State University and Agricultural Mechanical College: Baton Rouge, LA, USA, 2021. [Google Scholar]
- Yang, Q.Q.; Liu, S.W.; He, C.; Yu, X.P. Distribution and the origin of invasive apple snails, Pomacea canaliculata and P. maculata (Gastropoda: Ampullariidae) in China. Sci. Rep. 2018, 8, 1185. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Matsukura, K. Seasonal changes in cold hardiness of the invasive freshwater apple snail, Pomacea canaliculata (Lamarck)(Gastropoda: Ampullariidae). Malacologia 2007, 49, 383–392. [Google Scholar] [CrossRef]
- Seuffert, M.E.; Martín, P.R. Influence of temperature, size and sex on aerial respiration of Pomacea canaliculata (Gastropoda: Ampullariidae) from southern Pampas, Argentina. Malacologia 2009, 51, 191–200. [Google Scholar] [CrossRef]
- Seuffert, M.E.; Burela, S.; Martín, P.R. Influence of water temperature on the activity of the freshwater snail Pomacea canaliculata (Caenogastropoda: Ampullariidae) at its southernmost limit (Southern Pampas, Argentina). J. Therm. Biol. 2010, 35, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Lith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar]
- Cayuela, L.; Golicher, D.; Newton, A.; Kolb, M.; De Alburquerque, F.; Arets, E.; Alkemade, J.; Pérez, A. Species distribution modeling in the tropics: Problems, potentialities, and the role of biological data for effective species conservation. Trop. Conserv. Sci. 2009, 2, 319–352. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Rome, Q.; Villemant, C.; Courchamp, F. Can species distribution models really predict the expansion of invasive species? PLoS ONE 2018, 13, e0193085. [Google Scholar] [CrossRef]
- Guo, C.; Lek, S.; Ye, S.; Li, W.; Liu, J.; Li, Z. Uncertainty in ensemble modelling of large-scale species distribution: Effects from species characteristics and model techniques. Ecol. Model. 2015, 306, 67–75. [Google Scholar] [CrossRef]
- Loh, W. Classification and regression trees. Wires. Data. Min. Knowl. 2011, 1, 14–23. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Chen, T.; Guestrin, C. Xgboost: A scalable tree boosting system. In Proceedings of the 22nd Acm Sigkdd International Conference on Knowledge Discovery and Data Mining, San Francisco, CA, USA, 13–17 August 2016; pp. 785–794. [Google Scholar]
- Nelder, J.A.; Wedderburn, R.W. Generalized linear models. J. R. Stat. Soc. Ser. A Stat. Soc. 1972, 135, 370–384. [Google Scholar] [CrossRef]
- Hastie, T.J. Generalized additive models. In Statistical Models in S; Routledge: Oxfordshire, UK, 2017; pp. 249–307. [Google Scholar]
- Berger, A.; Della Pietra, S.A.; Della Pietra, V.J. A maximum entropy approach to natural language processing. Comput. Linguist. 1996, 22, 39–71. [Google Scholar]
- Lomba, A.; Pellissier, L.; Randin, C.; Vicente, J.; Moreira, F.; Honrado, J.; Guisan, A. Overcoming the rare species modelling paradox: A novel hierarchical framework applied to an Iberian endemic plant. Biol. Conserv. 2010, 143, 2647–2657. [Google Scholar] [CrossRef]
- Jones-Farrand, D.T.; Fearer, T.M.; Thogmartin, W.E.; Thompson, F.R., III; Nelson, M.D.; Tirpak, J.M. Comparison of statistical and theoretical habitat models for conservation planning: The benefit of ensemble prediction. Ecol. Appl. 2011, 21, 2269–2282. [Google Scholar] [CrossRef] [PubMed]
- Dormann, C.F.; Calabrese, J.M.; Guillera-Arroita, G.; Matechou, E.; Bahn, V.; Bartoń, K.; Beale, C.M.; Ciuti, S.; Elith, J.; Gerstner, K. Model averaging in ecology: A review of Bayesian, information-theoretic, and tactical approaches for predictive inference. Ecol. Monogr. 2018, 88, 485–504. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A review of evidence about use and performance of species distribution modelling ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- Park, C.; Byeon, J.; Park, J.; Lee, S.; Park, B.; Kim, J.; Woo, H. Impact of climate change on species distribution of Picea jezoensis Carrière in Baekdudaegan using ensemble modeling. Sens. Mater. 2022, 34, 4629–4637. [Google Scholar] [CrossRef]
- Huang, D.; An, Q.; Huang, S.; Tan, G.; Quan, H.; Chen, Y.; Zhou, J.; Liao, H. Biomod2 modeling for predicting the potentialecological distribution of three Fritillaria species under climate change. Sci. Rep. 2023, 13, 18801. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Cui, X.; Sun, J.; Li, T.; Wang, Q.; Ye, X.; Fan, B. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models. Ecol. Indic. 2021, 132, 108256. [Google Scholar] [CrossRef]
- Yin, Y.; He, Q.; Pan, X.; Liu, Q.; Wu, Y.; Li, X. Historical invasion, expansion process and harm investigation of Pomacea canaliculata in China. Biology 2022, 11, 110. [Google Scholar]
- Zhang, H.; Luo, D.; Mu, X.; Xu, M.; Wei, H.; Luo, J.; Zhang, J.; Hu, Y. Predicting the potential suitable distribution area of the apple snail Pomacea canaliculata in China based on multiple ecological niche models. Chin. J. Appl. Ecol. 2016, 27, 1277–1284. [Google Scholar]
- Wang, Y.; Zheng, P.; Pan, W. Predicting the potential suitable distribution area of Pomacea canliculate in China based on the GARP ecological niche modeling. J. Fujian Agric. For. Univ. 2018, 47, 21–25. [Google Scholar]
- Zhao, Z.; Xiao, N.; Shen, M.; Li, J. Comparison between optimized MaxEnt and random forest modeling in predicting potential distribution: A case study with Quasipaa boulengeri in China. Sci. Total Environ. 2022, 842, 156867. [Google Scholar] [CrossRef] [PubMed]
- Crego, R.D.; Stabach, J.A.; Connette, G. Implementation of species distribution models in Google Earth Engine. Divers. Distrib. 2022, 28, 904–916. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Rashid, W.; Shi, J.; Rahim, I.; Qasim, M.; Baloch, M.N.; Bohnett, E.; Yang, F.; Khan, I.; Ahmad, B. Modelling potential distribution of snow Leopards in pamir, northern Pakistan: Implications for human–snow leopard conflicts. Sustainability 2021, 13, 13229. [Google Scholar] [CrossRef]
- Fawcett, T. An introduction to ROC analysis. Pattern Recogn. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]
- Huang, J.; Ling, C.X. Using AUC and accuracy in evaluating learning algorithms. IEEE Trans. Knowl. Data Eng. 2005, 17, 299–310. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Brotons, L.; Thuiller, W.; Araújo, M.B.; Hirzel, A.H. Presence-absence versus presence-only modelling methods for predicting bird habitat suitability. Ecography 2004, 27, 437–448. [Google Scholar] [CrossRef]
- MacLeod, C.D.; Mandleberg, L.; Schweder, C.; Bannon, S.M.; Pierce, G.J. A comparison of approaches for modelling the occurrence of marine animals. In Essential Fish Habitat Mapping in the Mediterranean; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008; pp. 21–32. [Google Scholar]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Breiner, F.T.; Guisan, A.; Bergamini, A.; Nobis, M.P. Overcoming limitations of modelling rare species by using ensembles of small models. Methods Ecol. Evol. 2015, 6, 1210–1218. [Google Scholar] [CrossRef]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 2009, 15, 59–69. [Google Scholar] [CrossRef]
- Crimmins, S.M.; Dobrowski, S.Z.; Mynsberge, A.R. Evaluating ensemble forecasts of plant species distributions under climate change. Ecol. Model. 2013, 266, 126–130. [Google Scholar] [CrossRef]
- Deng, X.; Liu, D.; Guo, X.; Tian, Z.; Yang, L.; Gu, R.; Huang, Z.; Li, Y.; Liu, S.; Xiao, W. Temporal and spatial distribution of Pomacea canaliculata in Er’hai Lake catchment. Asian J. Ecotox. 2018, 13, 134–142. [Google Scholar]
- Lv, S.; Zhang, Y.; Steinmann, P.; Yang, G.; Yang, K.; Zhou, X.; Utzinger, J. The emergence of angiostrongyliasis in the People’s Republic of China: The interplay between invasive snails, climate change and transmission dynamics. Freshw. Biol. 2011, 56, 717–734. [Google Scholar] [CrossRef]
- Qin, Z.; Zhang, J.; Yao, F.; Liu, J.; Shi, Z.; Zhao, B.; Guo, J. Niche conservatism and geographical range expansion of Pomacea canaliculata and Pomacea maculata in non-native United States and China. Biol. Invasions 2023, 25, 3391–3405. [Google Scholar] [CrossRef]
- Yoshida, K.; Hoshikawa, K.; Wada, T.; Yusa, Y. Life cycle of the apple snail Pomacea canaliculata (Caenogastropoda: Ampullariidae) inhabiting Japanese paddy fields. Appl. Entomol. Zool. 2009, 44, 465–474. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Z.; Wang, Z.; Peng, Y. A comparative transcriptomics approach to analyzing the differences in cold resistance in Pomacea canaliculata between Guangdong and Hunan. J. Immunol. Res. 2020, 2020, 8025140. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Zhang, T.; An, W.; Wang, Z.; Li, C. Predicting the Potential Geographic Distribution of Invasive Freshwater Apple Snail Pomacea canaliculate (Lamarck, 1819) under Climate Change Based on Biomod2. Agronomy 2024, 14, 650. https://doi.org/10.3390/agronomy14040650
Wang T, Zhang T, An W, Wang Z, Li C. Predicting the Potential Geographic Distribution of Invasive Freshwater Apple Snail Pomacea canaliculate (Lamarck, 1819) under Climate Change Based on Biomod2. Agronomy. 2024; 14(4):650. https://doi.org/10.3390/agronomy14040650
Chicago/Turabian StyleWang, Tao, Tingjia Zhang, Weibin An, Zailing Wang, and Chuanren Li. 2024. "Predicting the Potential Geographic Distribution of Invasive Freshwater Apple Snail Pomacea canaliculate (Lamarck, 1819) under Climate Change Based on Biomod2" Agronomy 14, no. 4: 650. https://doi.org/10.3390/agronomy14040650