
Shoot Development in Members of an Ancient Aquatic Angiosperm Lineage, Ceratophyllaceae: A New Interpretation Facilitates Comparisons with Chloranthaceae
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Ceratophyllum submersum
3.1.1. Young Primary Shoot of Seed Embryo and Initiation of Its Phyllotaxis
3.1.2. Initiation of Phyllotaxis in Lateral Shoots
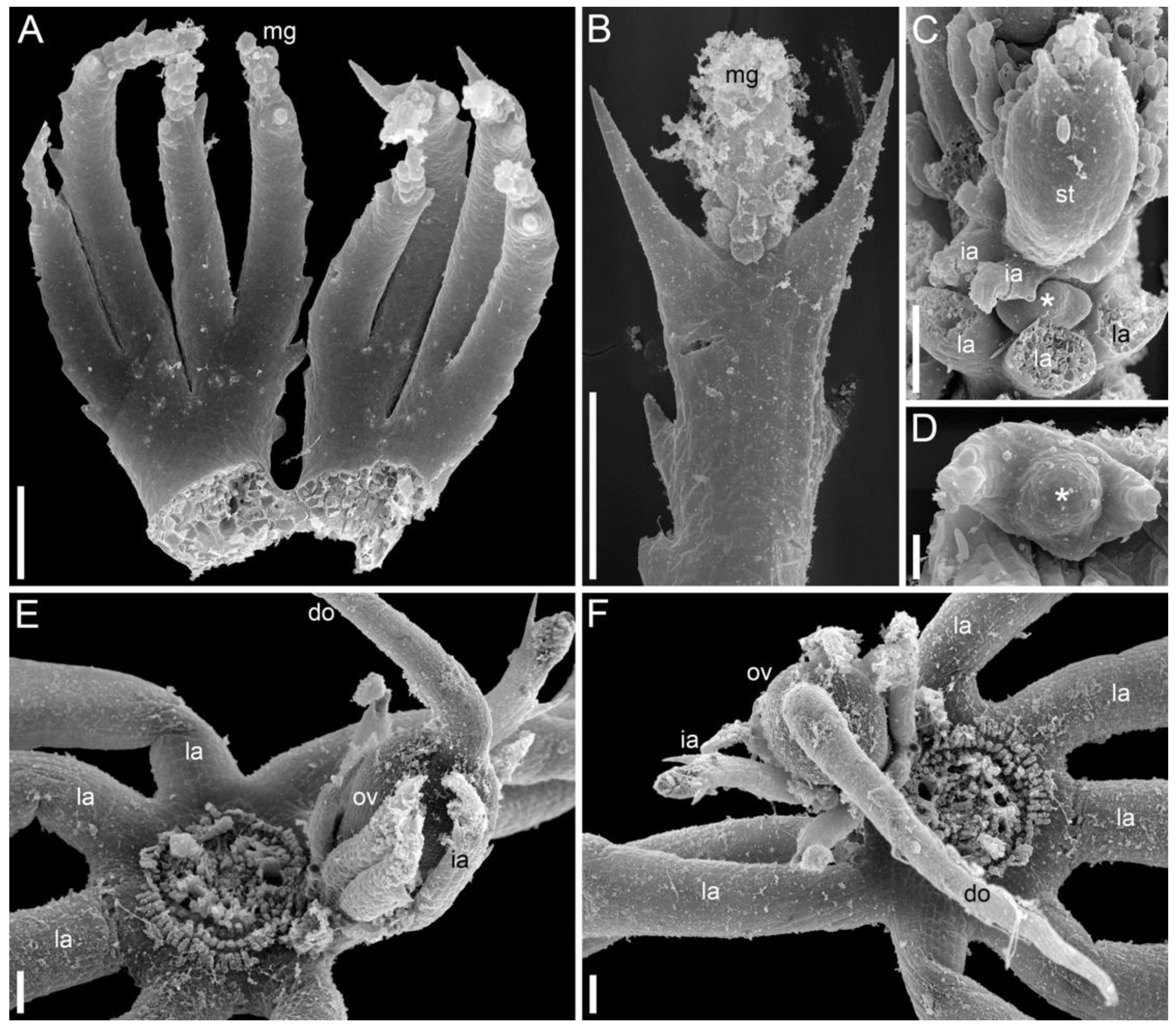
3.1.3. Morphology of Mature Shoots
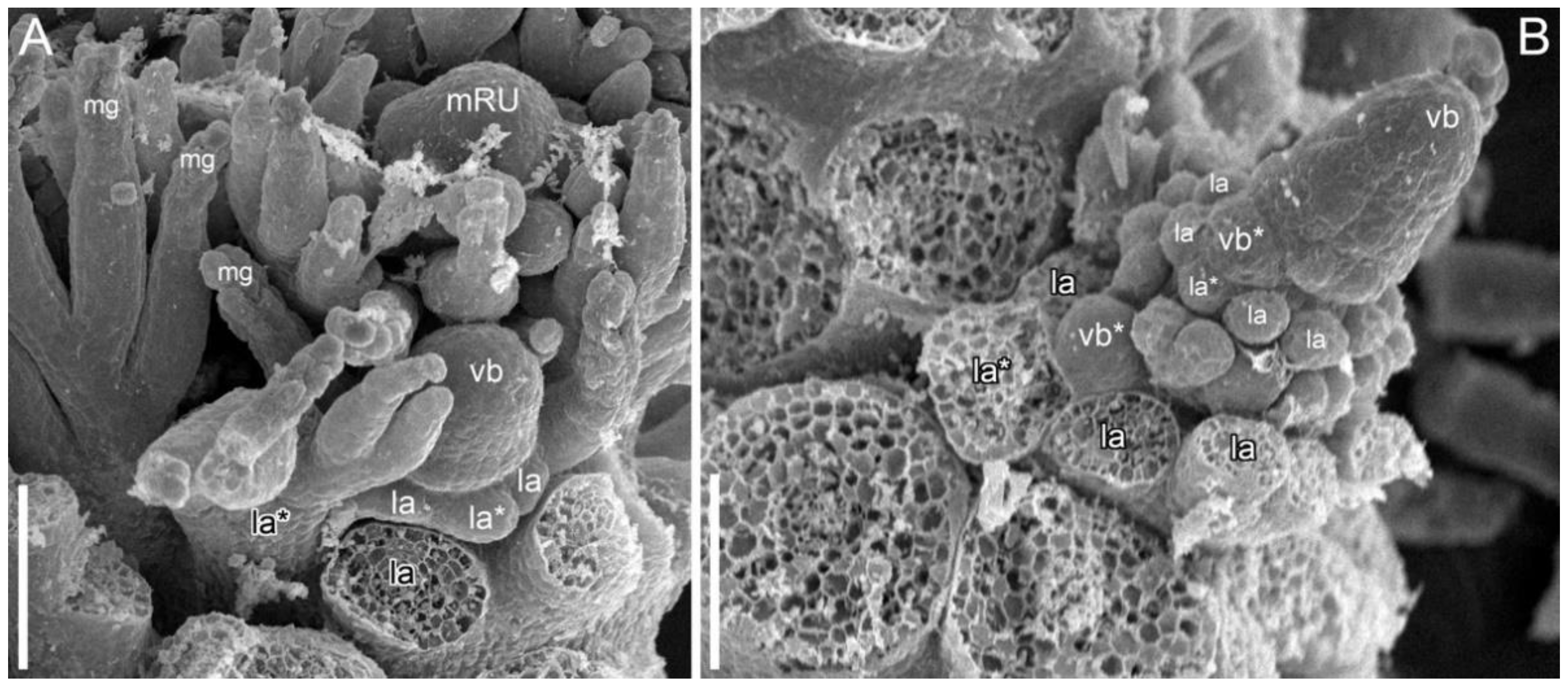
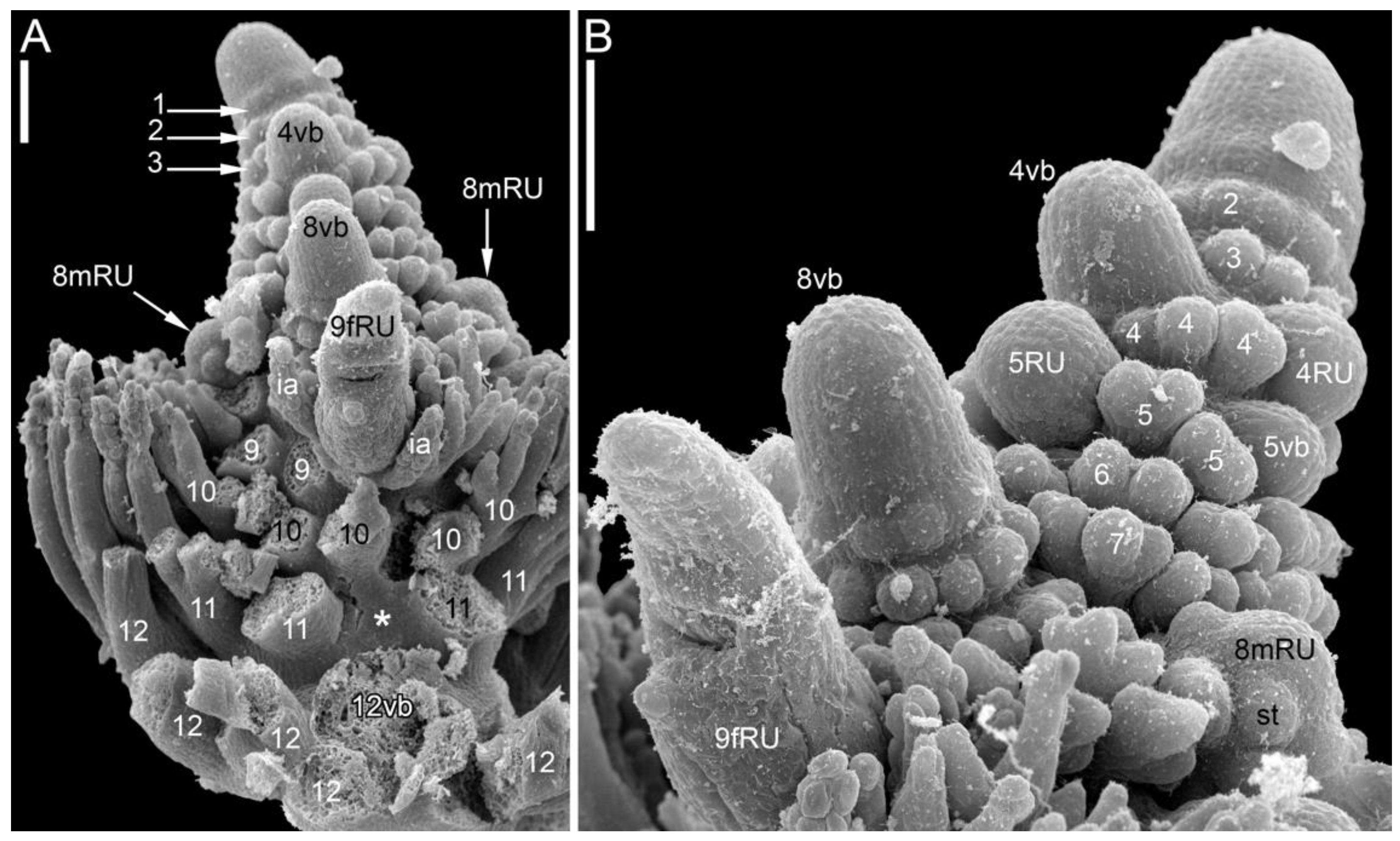
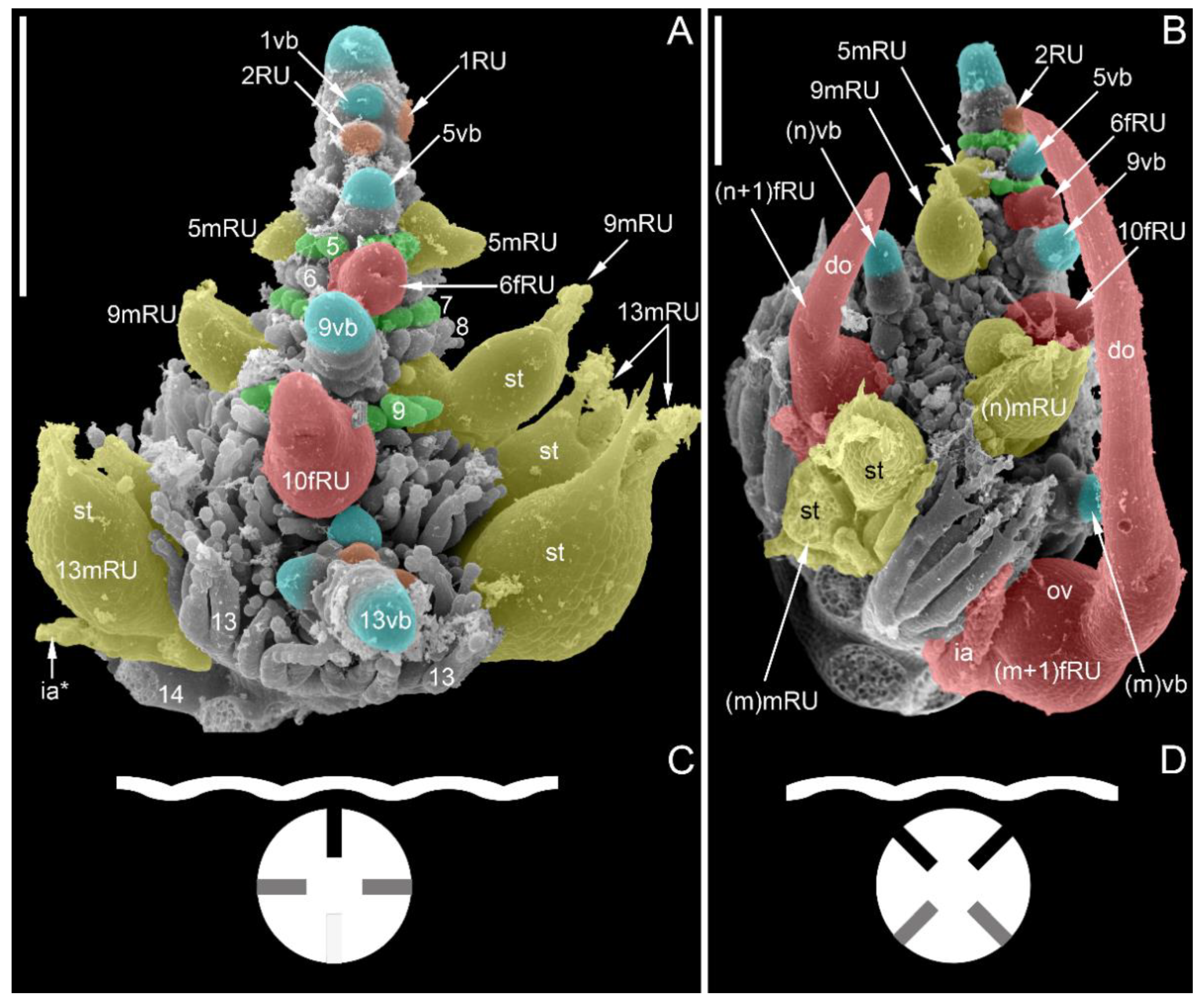
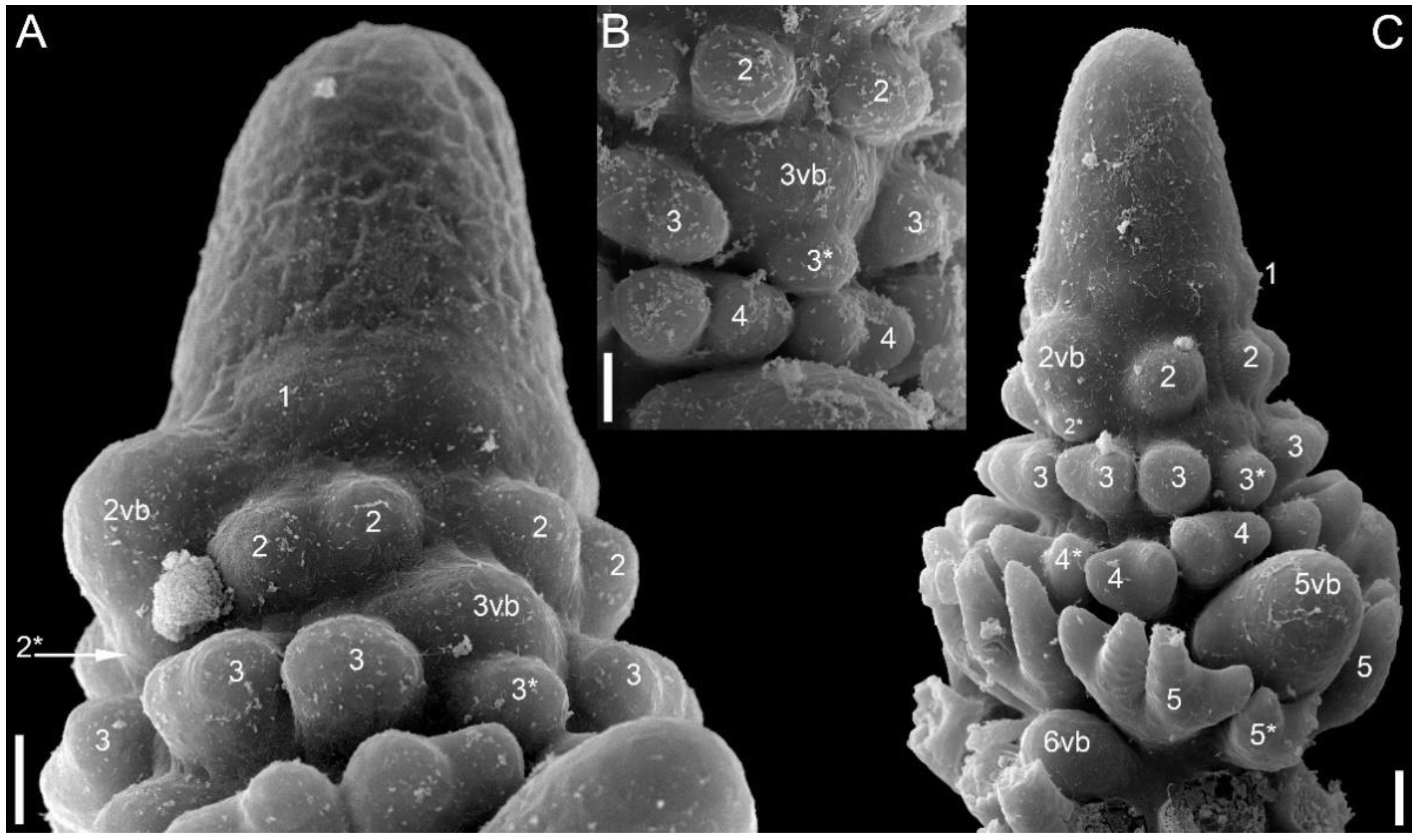
3.1.4. SEM-Based Description of Young Shoot Tips
3.2. Ceratophyllum tanaiticum
3.3. Ceratophyllum demersum
4. Discussion
- (1)
- Raynal-Roques (1981) [22] described occasional occurrence of as many as four male reproductive units per node in Ceratophyllum, with two units larger than two others (Figure 2C). Raynal-Roques [22] interpreted all four male reproductive units as accessory collateral buds in the axil of one and the same leaf in the framework of the concept of dorsiventral distichy. Iwamoto et al. [25], in the framework of the decussate hypothesis, concluded that each of two leaves has a pair of collateral accessory buds. Our hypothesis implies that each node should not possess more than two reproductive units (Figure 2F). The following arguments can be used to defend our hypothesis against the problematic observation of Raynal-Roques [22]. No photographs of the nodes with four reproductive unts are available, only line drawings. None of other studies revealed this pattern. The male units are situated in closely spaced pairs, with one unit younger than the other. We speculate that the younger units may represent secondary branches of the older units. Branching of female reproductive units was documented in detail by Aboy [40]. Testing this hypothesis requires a developmental analysis of material similar to what is illustrated by Raynal-Roques [22].
- (2)
- Our hypothesis implies that, in some instances, out of two serial buds only one is present. When only one bud is present, this can be either a vegetative branch of a reproductive unit. Therefore, in certain instances, only an accessory bud is present and the principal bud is absent. We do not consider this point too problematic, because completely the same problem occurs with the collateral bud hypothesis with respect to shoots of Ceratophyllum tanaiticum possessing only one orthostichy of lateral axes (Figure 7).
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gatsuk, L.E. Gemmaxillary plants and a system of subordinated units of their shoot body. Bull. Mosc. Soc. Nat. Biol. Ser. 1974, 79, 100–113. [Google Scholar]
- Doyle, J.A. Phylogenetic analyses and morphological innovations in land plants. Annu. Plant Rev. 2013, 45, 5. [Google Scholar]
- Bierhorst, D.W. Morphology of Vascular Plants; Macmillan: New York, NY, USA, 1971. [Google Scholar]
- Bell, A.D.; Bryan, A. Plant Form: An Illustrated Guide to Flowering Plant Morphology; Timber Press: London, UK; Portland, OR, USA, 2008. [Google Scholar]
- Weberling, F. Morphology of Flowers and Inflorescences; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Baum, D.A.; Day, C.D. Cryptic bracts exposed: Insights into the regulation of leaf expansion. Dev. Cell 2004, 6, 318–319. [Google Scholar] [CrossRef] [Green Version]
- Penin, A.A. Bract reduction in Cruciferae: Possible genetic mechanisms and evolution. Wulfenia 2008, 15, 63–73. [Google Scholar]
- Endress, P.K.; Doyle, J.A. Reconstructing the ancestral angiosperm flower and its initial specializations. Am. J. Bot. 2009, 96, 22–66. [Google Scholar] [CrossRef]
- Remizowa, M.V.; Sokoloff, D.D.; Rudall, P.J. Patterns of bract reduction in racemose inflorescences of early-divergent monocots. In Early Events in Monocot Evolution; Wilkin, P., Mayo, S.J., Eds.; Cambridge University Press: Cambridge, UK, 2013; pp. 185–207. ISBN 978-1-139-00295-0. [Google Scholar]
- Stevenson, D.W. Observations on vegetative branching in cycads. Int. J. Plant Sci. 2020, 181, 564–580. [Google Scholar] [CrossRef]
- Raciborski, M. Die Morphologie der Cabombeen und Nymphaeaceen. Flora 1894, 78, 244–279. [Google Scholar]
- Raciborski, M. Beiträge zur Kentniss der Cabombeen und Nymphaeaceen. Flora 1894, 79, 92–108. [Google Scholar]
- Cutter, E. Studies of morphogenesis in the Nymphaeaceae. I. Introduction: Some aspects of the morphology of Nuphar lutea (L.) Sm. and Nymphaea alba L. Phytomorphology 1957, 7, 45–56. [Google Scholar]
- Cutter, E.G. The inception and distribution of flowers in the Nymphaeaceae. Proc. Linn. Soc. Lond. 1961, 172, 93–100. [Google Scholar] [CrossRef]
- Richardson, F.C. Morphological Studies of the Nymphaeaceae. IV. Structure and Development of the Flower of Brasenia schreberi Gmel; University of California Press: Berkeley, CA, USA, 1969; Volume 47, pp. 1–101. [Google Scholar]
- Moseley, M.F. Morphological studies of the Nymphaeaceae. VI. Development of the flower of Nuphar. Phytomorphology 1971, 21, 253–283. [Google Scholar]
- Schneider, E.L.; Tucker, S.C.; Williamson, P.S. Floral development in the Nymphaeales. Int. J. Plant Sci. 2003, 164, S279–S292. [Google Scholar] [CrossRef]
- Grob, V.; Moline, P.; Pfeifer, E.; Novelo, A.R.; Rutishauser, R. Developmental morphology of branching flowers in Nymphaea prolifera. J. Plant Res. 2006, 119, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Rudall, P.J.; Sokoloff, D.D.; Remizowa, M.V.; Conran, J.G.; Davis, J.I.; Macfarlane, T.D.; Stevenson, D.W. Morphology of Hydatellaceae, an anomalous aquatic family recently recognized as an early-divergent angiosperm lineage. Am. J. Bot. 2007, 94, 1073–1092. [Google Scholar] [CrossRef] [Green Version]
- Sokoloff, D.D.; Remizowa, M.V.; Briggs, B.G.; Rudall, P.J. Shoot architecture and branching pattern in perennial Hydatellaceae (Nymphaeales). Int. J. Plant Sci. 2009, 170, 869–884. [Google Scholar] [CrossRef]
- El, E.S.; Remizowa, M.V.; Sokoloff, D.D. Developmental flower and rhizome morphology in Nuphar (Nymphaeales): An interplay of chaos and stability. Front. Cell Dev. Biol. 2020, 8, 303. [Google Scholar] [CrossRef]
- Raynal-Roques, A. Contribution à L’étude Biomorphologique des Angiospermes Aquatiques Tropicales: Essai D’analyse De l’évolution. Ph.D. Thesis, Université des Sciences et Techniques du Languedoc, Montpellier, France, 1981. [Google Scholar]
- Rutishauser, R.; Sattler, R. Complementarity and heuristic value of contrasting models in structural botany. II. Case study on leaf whorls: Equisetum and Ceratophyllum. Bot. Jahrb. Syst. 1987, 109, 227–255. [Google Scholar]
- Rutishauser, R. Polymerous leaf whorls in vascular plants: Developmental morphology and fuzziness of organ identities. Int. J. Plant Sci. 1999, 160, S81–S103. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, A.; Izumidate, R.; Ronse De Craene, L.P. Floral anatomy and vegetative development in Ceratophyllum demersum: A morphological picture of an “unsolved” plant. Am. J. Bot. 2015, 102, 1578–1589. [Google Scholar] [CrossRef] [Green Version]
- Gitzendanner, M.A.; Soltis, P.S.; Wong, G.K.-S.; Ruhfel, B.R.; Soltis, D.E. Plastid phylogenomic analysis of green plants: A billion years of evolutionary history. Am. J. Bot. 2018, 105, 291–301. [Google Scholar] [CrossRef]
- Leebens-Mack, J.H.; Barker, M.S.; Carpenter, E.J.; Deyholos, M.K.; Gitzendanner, M.A.; Graham, S.W.; Grosse, I.; Li, Z.; Melkonian, M.; Mirarab, S.; et al. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef] [Green Version]
- Albert, V.A.; Renner, T. Aquatic angiosperm ambiguities answered. Nat. Plants 2020, 6, 181–183. [Google Scholar] [CrossRef] [PubMed]
- Mavrodiev, E.V.; Williams, D.M.; Ebach, M.C.; Mavrodieva, A.E. Fassettia, a new North American genus of family Ceratophyllaceae: Evidence based on cladistic analyses of current molecular data of Ceratophyllum. Aust. Syst. Bot. 2021, 34, 431–437. [Google Scholar] [CrossRef]
- Szalontai, B.; Stranczinger, S.; Mesterházy, A.; Scribailo, R.W.; Les, D.H.; Efremov, A.N.; Jacono, C.C.; Kipriyanova, L.M.; Kaushik, K.; Laktionov, A.P.; et al. Molecular phylogenetic analysis of Ceratophyllum L. taxa: A new perspective. Bot. J. Linn. Soc. 2018, 188, 161–172. [Google Scholar] [CrossRef]
- Guo, X.; Fang, D.; Sahu, S.K.; Yang, S.; Guang, X.; Folk, R.; Smith, S.A.; Chanderbali, A.S.; Chen, S.; Liu, M.; et al. Chloranthus genome provides insights into the early diversification of angiosperms. Nat. Commun. 2021, 12, 6930. [Google Scholar] [CrossRef]
- Doyle, J.A.; Endress, P.K. Phylogenetic analyses of Cretaceous fossils related to Chloranthaceae and their evolutionary implications. Bot. Rev. 2018, 84, 156–202. [Google Scholar] [CrossRef] [Green Version]
- Kvaček, J.; Doyle, J.A.; Endress, P.K.; Daviero-Gomez, V.; Gomez, B.; Tekleva, M. Pseudoasterophyllites cretaceus from the Cenomanian (Cretaceous) of the Czech Republic: A possible link between Chloranthaceae and Ceratophyllum. Taxon 2016, 65, 1345–1373. [Google Scholar] [CrossRef] [Green Version]
- Gomez, B.; Daviero-Gomez, V.; Coiffard, C.; Barral, A.; Martín-Closas, C.; Dilcher, D.L. Montsechia vidalii from the Barremian of Spain, the earliest known submerged aquatic angiosperm, and its systematic relationship to Ceratophyllum. Taxon 2020, 69, 1273–1292. [Google Scholar] [CrossRef]
- Antonov, A.S.; Troitsky, A.V.; Samigullin, T.K.; Bobrova, V.K.; Valiejo-Roman, K.M.; Martin, W. Early events in the evolution of angiosperms deduced from Cp RDNA ITS 2–4 sequence comparisons. In Proceedings of the International Symposium on the Family Magnoliaceae; Liu, Y.-H., Fan, H.-M., Chen, Z.-Y., Wu, Q.-G., Zeng, Q.-W., Eds.; Science Press: Beijing, China, 2000; pp. 210–214. [Google Scholar]
- Moore, M.J.; Hassan, N.; Gitzendanner, M.A.; Bruenn, R.A.; Croley, M.; Vandeventer, A.; Horn, J.W.; Dhingra, A.; Brockington, S.F.; Latvis, M.; et al. Phylogenetic analysis of the plastid inverted repeat for 244 species: Insights into deeper-level angiosperm relationships from a long, slowly evolving sequence region. Int. J. Plant Sci. 2011, 172, 541–558. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Zhang, Q.; Sun, R.; Kong, H.; Zhang, N.; Ma, H. Resolution of deep angiosperm phylogeny using conserved nuclear genes and estimates of early divergence times. Nat. Commun. 2014, 5, 4956. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.-Y.; Dong, S.-S.; Wang, M.-Q.; Song, T.-Q.; Zhou, G.-C.; Li, Z.; de Peer, Y.V.; Shao, Z.-Q.; Wang, W.; Chen, M.; et al. Mitochondrial genes from 18 angiosperms fill sampling gaps for phylogenomic inferences of the early diversification of flowering plants. J. Syst. Evol. 2021; Early View. [Google Scholar] [CrossRef]
- Strasburger, E. Ein Beitrag zur Kenntnis von Ceratophyllum submersum und phylogenetische Erörterungen. Jahrb. Wiss. Bot. 1902, 37, 477–526. [Google Scholar]
- Aboy, H.E. A Study of the Anatomy and Morphology of Ceratophyllum demersum. Master’s Thesis, Cornell University, Ithaca, NY, USA, 1936. [Google Scholar]
- Sehgal, A.; Ram, H.Y.M. Comparative developmental morphology of two populations of Ceratophyllum L. (Ceratophyllaceae) and their taxonomy. Bot. J. Linn. Soc. 1981, 82, 343–356. [Google Scholar] [CrossRef]
- Shamrov, I.I. Anthecological investigation of three species of the genus Ceratophyllum (Ceratophyllaceae). Bot. Zhurnal 1983, 68, 1357–1366. [Google Scholar]
- Les, D.H. Systematics and Evolution of Ceratophyllum L. (Ceratophyllaceae): A Monograph (Taxonomy, Flavonoids, Aquatic plants, Phytogeography, Variation). Ph.D. Thesis, The Ohio State University, Ann Arbor, MI, USA, 1986. [Google Scholar]
- Endress, P.K. Evolutionary aspects of the floral structure in Ceratophyllum. Plant Syst. Evol. 1994, 8, 175–183. [Google Scholar] [CrossRef]
- Endress, P.K. The flowers in extant basal angiosperms and inferences on ancestral flowers. Int. J. Plant Sci. 2001, 162, 1111–1140. [Google Scholar] [CrossRef]
- Iwamoto, A.; Shimizu, A.; Ohba, H. Floral development and phyllotactic variation in Ceratophyllum demersum (Ceratophyllaceae). Am. J. Bot. 2003, 90, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Endress, P.K.; Doyle, J.A. Ancestral traits and specializations in the flowers of the basal grade of living angiosperms. Taxon 2015, 64, 1093–1116. [Google Scholar] [CrossRef]
- Schaeppi, H. Untersuchungen über die Blattentwicklung bei Ceratophyllum, Cabomba und Limnophila. Planta 1935, 24, 755–769. [Google Scholar] [CrossRef]
- Shamrov, I.I. Morphological nature of gynoecium and fruit in Ceratophyllum (Ceratophyllaceae). Bot. Zhurnal 2009, 94, 938–961. [Google Scholar]
- Sokoloff, D.D.; El, E.S.; Pechenyuk, E.V.; Carrive, L.; Nadot, S.; Rudall, P.J.; Remizowa, M.V. Refined interpretation of the female flower in Ceratophyllum sheds fresh light on gynoecium evolution in angiosperms. Front. Cell Dev. Biol. 2022, 10, 868352. [Google Scholar] [CrossRef]
- Endress, P.K. Structure and relationships of basal relictual angiosperms. Aust. Syst. Bot. 2004, 17, 343–366. [Google Scholar] [CrossRef]
- Takhtajan, A. Systema et Phylogenia Magnoliophytorum; Moscow & Leningrad: Nauka, Russia, 1966. (In Russian) [Google Scholar]
- Cronquist, A. An Integrated System of Classification of Flowering Plants; Columbia University Press: New York, NY, USA, 1981. [Google Scholar]
- Goebel, K.E. Über die Verzweigung dorsiventraler Sprosse. Arb. Bot. Inst. Wuerzb. 1882, 2, 353–436. [Google Scholar]
- Schneider, O.H. Zur Kenntnis dorsiventraler Blütenstände in der Familie der Papilionaceen. Bot. Arch. 1925, 11, 26–39. [Google Scholar]
- Charlton, W.A. Pendulum symmetry. In Symmetry in Plants; Barabé, D., Jean, R.V., Eds.; World Scientific: Singapore, 1998; pp. 61–87. ISBN 978-981-02-2621-3. [Google Scholar]
- Loiseau, J.E.; Grangeon, D. Variations phyllotaxiques chez Ceratophyllum demersum L. et Hippuris vulgaris L. Bull. Soc. Bot. Fr. 1963, 110, 76–91. [Google Scholar] [CrossRef] [Green Version]
- Serebryakova, T.I. Shoot Morphogenesis and the Evolution of Life Forms in Grasses; Nauka: Moscow, Russia, 1971. [Google Scholar]
- Endress, P.K. Disentangling confusions in inflorescence morphology: Patterns and diversity of reproductive shoot ramification in angiosperms. J. Syst. Evol. 2010, 48, 225–239. [Google Scholar] [CrossRef]
- Endress, P.K.; Lorence, D.H. Inflorescence structure in Laurales—Stable and flexible patterns. Int. J. Plant Sci. 2020, 181, 267–283. [Google Scholar] [CrossRef] [Green Version]
- Endress, P.K.; Igersheim, A. The reproductive structures of the basal angiosperm Amborella trichopoda (Amborellaceae). Int. J. Plant Sci. 2000, 161, S237–S248. [Google Scholar] [CrossRef]
- Skutch, A.F. Peculiarities in the structure of the stem, related to the leaf-sheath, in Hedyosmum. Ann. Bot. 1927, 41, 715–730. [Google Scholar] [CrossRef]
- Todzia, C.A. Four new species of Hedyosmum (Chloranthaceae) from South America. Syst. Bot. 1988, 13, 21–31. [Google Scholar] [CrossRef]
- Todzia, C.A. Chloranthaceae: Hedyosmum. Flora Neotrop. 1988, 48, 1–138. [Google Scholar]
- Todzia, C.A.; Wood, C.E. The genus Hedyosmum (Chloranthaceae) in the West Indies. J. Arnold Arbor. 1988, 69, 51–63. [Google Scholar] [CrossRef]
- Doria, M.G.; Pabón-Mora, N.; González, F. Reassessing inflorescence and floral morphology and development in Hedyosmum (Chloranthaceae). Int. J. Plant Sci. 2012, 173, 735–750. [Google Scholar] [CrossRef]
- Sender, L.M.; Doyle, J.A.; Upchurch, G.R., Jr.; Endress, P.K.; Villanueva-Amadoz, U.; Diez, J.B. Evidence on vegetative and inflorescence morphology of Chloranthaceae (Angiospermae) from the Early Cretaceous (middle–late Albian) of Spain. J. Systemat. Palaeontol. 2021, 18, 2015–2042. [Google Scholar] [CrossRef]
- Ascarina lucida—The University of Auckland. Available online: https://www.nzplants.auckland.ac.nz/en/about/seed-plants-flowering/chloranthaceae/ascarina-lucida.html (accessed on 14 March 2022).
- D’Arcy, W.G.; Liesner, R.L. Hedyosmum (Chloranthaceae) in Panama. Syst. Bot. 1981, 6, 74–86. [Google Scholar] [CrossRef]
- Dormer, K.J.; Cutter, E.G. On the arrangement of flowers on the rhizomes of some Nymphaeaceae. New Phytol. 1959, 58, 176–181. [Google Scholar] [CrossRef]

















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokoloff, D.D.; El, E.S.; Remizowa, M.V. Shoot Development in Members of an Ancient Aquatic Angiosperm Lineage, Ceratophyllaceae: A New Interpretation Facilitates Comparisons with Chloranthaceae. Symmetry 2022, 14, 1288. https://doi.org/10.3390/sym14071288
Sokoloff DD, El ES, Remizowa MV. Shoot Development in Members of an Ancient Aquatic Angiosperm Lineage, Ceratophyllaceae: A New Interpretation Facilitates Comparisons with Chloranthaceae. Symmetry. 2022; 14(7):1288. https://doi.org/10.3390/sym14071288
Chicago/Turabian StyleSokoloff, Dmitry D., Elena S. El, and Margarita V. Remizowa. 2022. "Shoot Development in Members of an Ancient Aquatic Angiosperm Lineage, Ceratophyllaceae: A New Interpretation Facilitates Comparisons with Chloranthaceae" Symmetry 14, no. 7: 1288. https://doi.org/10.3390/sym14071288