

Red Sea Atlas of Coral-Associated Bacteria Highlights Common Microbiome Members and Their Distribution across Environmental Gradients—A Systematic Review
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Red Sea Studies Survey
2.2. Coral-Associated Bacteria in the Red Sea
3. Results
3.1. Coral-Associated Bacteria Literature Survey
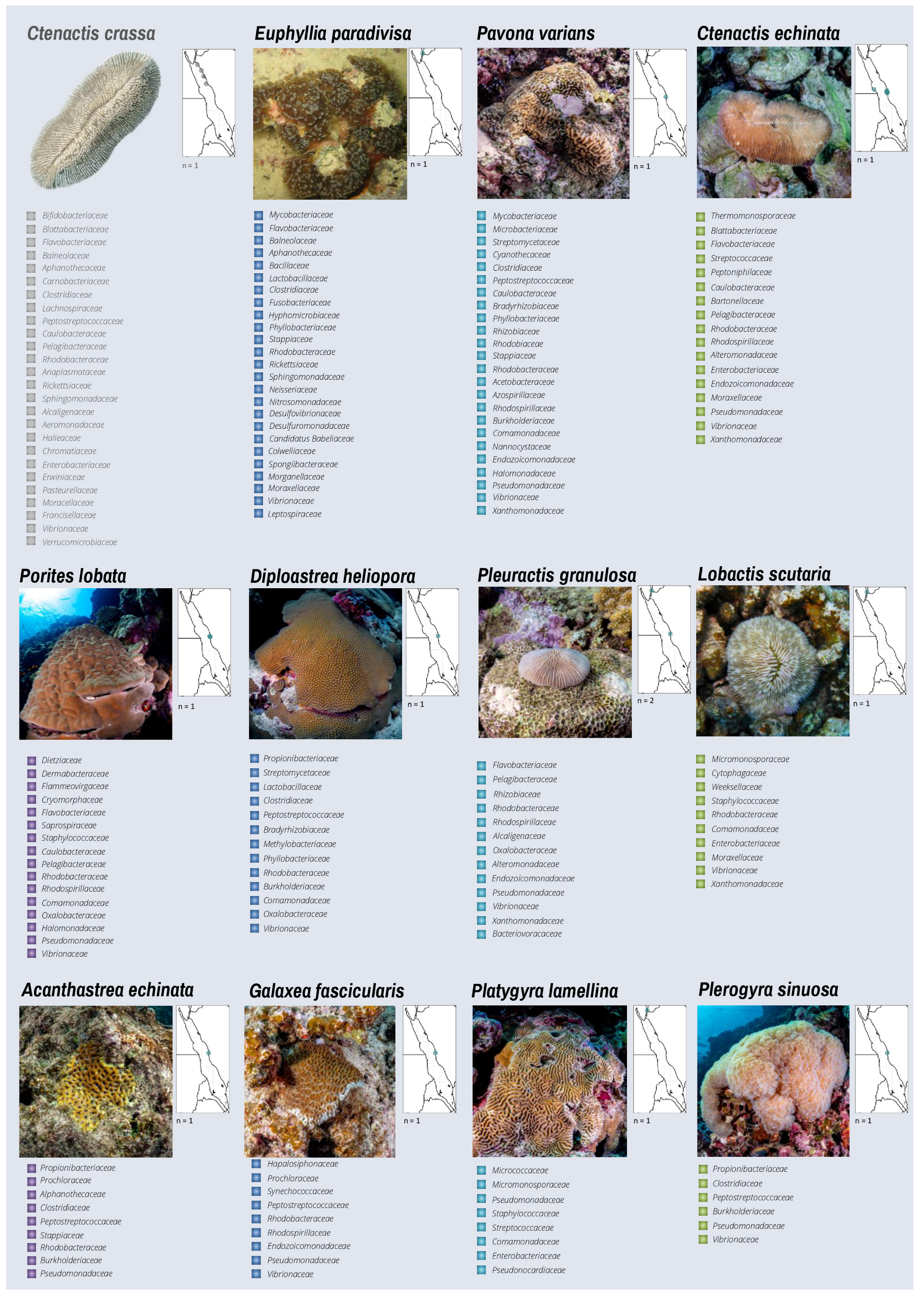
3.2. Bacterial Diversity of Red Sea Corals from Cultivation-Independent Techniques
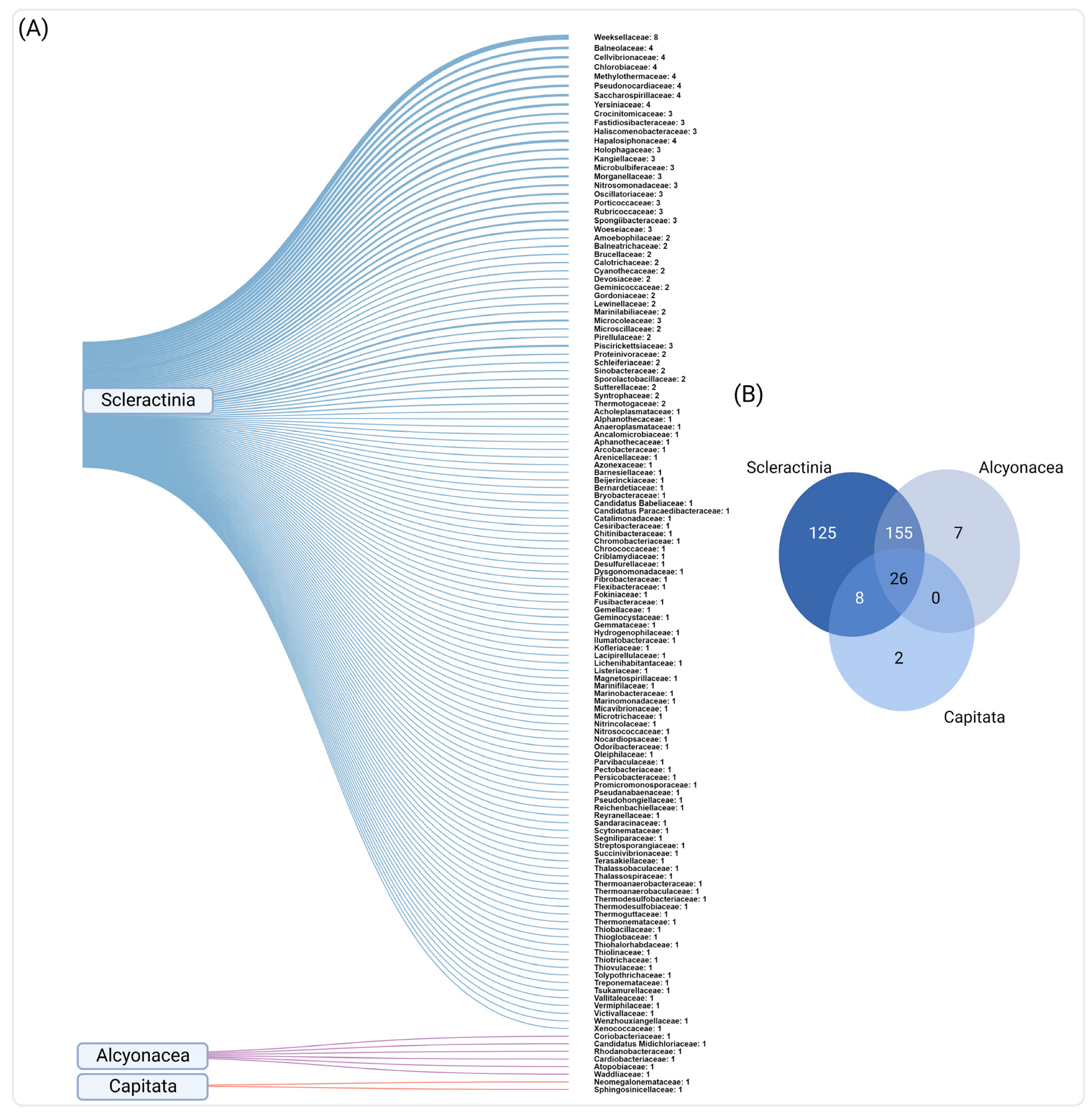
3.3. Common and Unique Bacteria in Red Sea Corals
3.4. Coral Bacteria across Red Sea Regions
3.5. Cultured Bacterial Diversity of Red Sea Corals
4. Discussion
4.1. Red Sea Coral Microbiome Survey
4.2. Culture-Independent Bacterial Diversity
4.3. Bacterial Diversity among Red Sea Regions
4.4. Cultured Bacterial Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rohwer, F.; Seguritan, V.; Azam, F.; Knowlton, N. Diversity and distribution of coral-associated bacteria. Mar. Ecol. Prog. Ser. 2002, 243, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Knowlton, N.; Rohwer, F. Multispecies microbial mutualisms on coral reefs: The host as a habitat. Am. Nat. 2003, 162, S51–S62. [Google Scholar] [CrossRef] [Green Version]
- Muscatine, L.; Goiran, C.; Land, L.; Jaubert, J.; Cuif, J.-P.; Allemand, D. Stable isotopes (Δ13C and Δ15N) of Organic matrix from coral skeleton. Proc. Natl. Acad. Sci. USA 2005, 102, 1525–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts. Curr. Biol. 2018, 28, 2570–2580.E6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, D.G.; Morrow, K.M.; Webster, N.S. Insights into the coral microbiome: Underpinning the health and resilience of reef ecosystems. Annu. Rev. Microbiol. 2016, 70, 317–340. [Google Scholar] [CrossRef] [PubMed]
- Krediet, C.J.; Ritchie, K.B.; Paul, V.J.; Teplitski, M. Coral-associated microorganisms and their roles in promoting coral health and thwarting diseases. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peixoto, R.S.; Rosado, P.M.; de Assis Leite, D.C.; Rosado, A.S.; Bourne, D.G. Beneficial Microorganisms for Corals (BMC): Proposed mechanisms for coral health and resilience. Front. Microbiol. 2017, 8, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pootakham, W.; Mhuantong, W.; Putchim, L.; Yoocha, T.; Sonthirod, C.; Kongkachana, W.; Sangsrakru, D.; Naktang, C.; Jomchai, N.; Thongtham, N.; et al. Dynamics of coral-associated microbiomes during a thermal bleaching event. MicrobiologyOpen 2018, 7, e00604. [Google Scholar] [CrossRef] [PubMed]
- Santoro, E.P.; Borges, R.M.; Espinoza, J.L.; Freire, M.; Messias, C.S.M.A.; Villela, H.D.M.; Pereira, L.M.; Vilela, C.L.S.; Rosado, J.G.; Cardoso, P.M.; et al. Coral microbiome manipulation elicits metabolic and genetic restructuring to mitigate heat stress and evade mortality. Sci. Adv. 2021, 7, eabg3088. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.R.; Rivera, H.E.; Closek, C.J.; Medina, M. Microbes in the coral holobiont: Partners through evolution, development, and ecological interactions. Front. Cell. Infect. Microbiol. 2015, 4, 176. [Google Scholar] [CrossRef]
- Fragoso ados Santos, H.; Duarte, G.A.S.; da Costa Rachid, C.T.; Chaloub, R.M.; Calderon, E.N.; de Barros Marangoni, L.F.; Bianchini, A.; Nudi, A.H.; do Carmo, F.L.; van Elsas, J.D.; et al. Impact of oil spills on coral reefs can be reduced by bioremediation using probiotic microbiota. Sci. Rep. 2015, 5, 18268. [Google Scholar] [CrossRef] [Green Version]
- Sweet, M.J.; Bulling, M.T. On the importance of the microbiome and pathobiome in coral health and disease. Front. Mar. Sci. 2017, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, M.; Seneca, F.O.; Yum, L.K.; Palumbi, S.R.; Voolstra, C.R. Bacterial community dynamics are linked to patterns of coral heat tolerance. Nat. Commun. 2017, 8, 14213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbins, S.J.; Singleton, C.M.; Chan, C.X.; Messer, L.F.; Geers, A.U.; Ying, H.; Baker, A.; Bell, S.C.; Morrow, K.M.; Ragan, M.A.; et al. A genomic view of the reef-building coral Porites Lutea and its microbial symbionts. Nat. Microbiol. 2019, 4, 2090–2100. [Google Scholar] [CrossRef] [PubMed]
- Rosado, P.M.; Leite, D.C.A.; Duarte, G.A.S.; Chaloub, R.M.; Jospin, G.; da Rocha, U.N.; Saraiva, J.P.; Dini-Andreote, F.; Eisen, J.A.; Bourne, D.G.; et al. Marine probiotics: Increasing coral resistance to bleaching through microbiome manipulation. ISME J. 2019, 13, 921–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweet, M.; Villela, H.; Keller-Costa, T.; Costa, R.; Romano, S.; Bourne, D.G.; Cárdenas, A.; Huggett, M.J.; Kerwin, A.H.; Kuek, F.; et al. Insights into the cultured bacterial fraction of corals. mSystems 2021, 6, e01249-20. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.P.; Villela, H.D.M.; Santos, H.F.; Duarte, G.A.S.; Ribeiro, J.R.; Ghizelini, A.M.; Vilela, C.L.S.; Rosado, P.M.; Fazolato, C.S.; Santoro, E.P.; et al. Multi-domain probiotic consortium as an alternative to chemical remediation of oil spills at coral reefs and adjacent sites. Microbiome 2021, 9, 118. [Google Scholar] [CrossRef]
- Ziegler, M.; Grupstra, C.G.B.; Barreto, M.M.; Eaton, M.; BaOmar, J.; Zubier, K.; Al-Sofyani, A.; Turki, A.J.; Ormond, R.; Voolstra, C.R. Coral bacterial community structure responds to environmental change in a host-specific manner. Nat. Commun. 2019, 10, 3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voolstra, C.R.; Ziegler, M. Adapting with Microbial Help: Microbiome flexibility facilitates rapid responses to environmental change. BioEssays 2020, 42, 2000004. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Agreda, A.; Leggat, W.; Bongaerts, P.; Herrera, C.; Ainsworth, T.D. Rethinking the coral microbiome: Simplicity exists within a diverse microbial biosphere. mBio 2018, 9, e00812-18. [Google Scholar] [CrossRef]
- Osman, E.O.; Suggett, D.J.; Voolstra, C.R.; Pettay, D.T.; Clark, D.R.; Pogoreutz, C.; Sampayo, E.M.; Warner, M.E.; Smith, D.J. Coral Microbiome composition along the northern Red Sea suggests high plasticity of bacterial and specificity of endosymbiotic dinoflagellate communities. Microbiome 2020, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Sweet, M.J.; Croquer, A.; Bythell, J.C. Bacterial assemblages differ between compartments within the coral holobiont. Coral Reefs 2011, 30, 39–52. [Google Scholar] [CrossRef]
- Pollock, F.J.; McMinds, R.; Smith, S.; Bourne, D.G.; Willis, B.L.; Medina, M.; Thurber, R.V.; Zaneveld, J.R. Coral-associated bacteria demonstrate phylosymbiosis and cophylogeny. Nat. Commun. 2018, 9, 4921. [Google Scholar] [CrossRef] [PubMed]
- Reshef, L.; Koren, O.; Loya, Y.; Zilber-Rosenberg, I.; Rosenberg, E. The coral probiotic hypothesis. Environ. Microbiol. 2006, 8, 2068–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogoreutz, C.; Rädecker, N.; Cárdenas, A.; Gärdes, A.; Wild, C.; Voolstra, C.R. Dominance of Endozoicomonas bacteria throughout coral bleaching and mortality suggests structural inflexibility of the Pocillopora verrucosa microbiome. Ecol. Evol. 2018, 8, 2240–2252. [Google Scholar] [CrossRef] [PubMed]
- DiBattista, J.D.; Roberts, M.B.; Bouwmeester, J.; Bowen, B.W.; Coker, D.J.; Lozano-Cortés, D.F.; Howard Choat, J.; Gaither, M.R.; Hobbs, J.-P.A.; Khalil, M.T.; et al. A review of contemporary patterns of endemism for shallow water reef fauna in the Red Sea. J. Biogeogr. 2016, 43, 423–439. [Google Scholar] [CrossRef] [Green Version]
- Voolstra, C.R.; Berumen, M.L. (Eds.) Coral Reefs of the Red Sea; Coral reefs of the world; Springer International Publishing: Cham, Switzerland, 2019; Volume 11, ISBN 978-3-030-05800-5. [Google Scholar]
- Osman, E.O.; Smith, D.J.; Ziegler, M.; Kürten, B.; Conrad, C.; El-Haddad, K.M.; Voolstra, C.R.; Suggett, D.J. Thermal refugia against coral bleaching throughout the northern Red Sea. Glob. Chang. Biol. 2018, 24, e474–e484. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.J. Red Sea; Elsevier: Amsterdam, The Netherlands, 1987; ISBN 978-1-4832-8599-3. [Google Scholar]
- Berumen, M.L.; Voolstra, C.R.; Daffonchio, D.; Agusti, S.; Aranda, M.; Irigoien, X.; Jones, B.H.; Morán, X.A.G.; Duarte, C.M. The Red Sea: Environmental gradients shape a natural laboratory in a nascent ocean. In Coral Reefs of the Red Sea; Voolstra, C.R., Berumen, M.L., Eds.; Coral reefs of the world; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–10. ISBN 978-3-030-05802-9. [Google Scholar]
- Robitzch, V.; Banguera-Hinestroza, E.; Sawall, Y.; Al-Sofyani, A.; Voolstra, C.R. Absence of genetic differentiation in the coral Pocillopora verrucosa along environmental gradients of the Saudi Arabian Red Sea. Front. Mar. Sci. 2015, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Arrigoni, R.; Berumen, M.L.; Chen, C.A.; Terraneo, T.I.; Baird, A.H.; Payri, C.; Benzoni, F. Species delimitation in the reef coral genera Echinophyllia and Oxypora (Scleractinia, Lobophylliidae) with a description of two new species. Mol. Phylogenet. Evol. 2016, 105, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Arrigoni, R.; Benzoni, F.; Terraneo, T.I.; Caragnano, A.; Berumen, M.L. Recent origin and semi-permeable species boundaries in the scleractinian coral genus Stylophora from the Red Sea. Sci. Rep. 2016, 6, 34612. [Google Scholar] [CrossRef]
- Terraneo, T.I.; Fusi, M.; Hume, B.C.C.; Arrigoni, R.; Voolstra, C.R.; Benzoni, F.; Forsman, Z.H.; Berumen, M.L. Environmental latitudinal gradients and host-specificity shape Symbiodiniaceae distribution in Red Sea Porites corals. J. Biogeogr. 2019, 46, 2323–2335. [Google Scholar] [CrossRef]
- van Oppen, M.J.H.; Blackall, L.L. Coral microbiome dynamics, functions and design in a changing world. Nat. Rev. Microbiol. 2019, 17, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Neave, M.J.; Michell, C.T.; Apprill, A.; Voolstra, C.R. Endozoicomonas genomes reveal functional adaptation and plasticity in bacterial strains symbiotically associated with diverse marine hosts. Sci. Rep. 2017, 7, 40579. [Google Scholar] [CrossRef] [Green Version]
- Neave, M.J.; Rachmawati, R.; Xun, L.; Michell, C.T.; Bourne, D.G.; Apprill, A.; Voolstra, C.R. Differential specificity between closely related corals and abundant Endozoicomonas endosymbionts across global scales. ISME J. 2017, 11, 186–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. BMJ 2009, 339, b2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kooperman, N.; Ben-Dov, E.; Kramarsky-Winter, E.; Barak, Z.; Kushmaro, A. Coral mucus-associated bacterial communities from natural and aquarium environments. FEMS Microbiol. Lett. 2007, 276, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raitsos, D.E.; Pradhan, Y.; Brewin, R.J.W.; Stenchikov, G.; Hoteit, I. Remote sensing the phytoplankton seasonal succession of the Red Sea. PLoS ONE 2013, 8, e64909. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.; Aylagas, E.; Villalobos, R.; Kattan, Y.; Berumen, M.; Pearman, J.K. Beyond the sisual: Using metabarcoding to characterize the hidden reef cryptobiome. Proc. R. Soc. B Biol. Sci. 2019, 286, 20182697. [Google Scholar] [CrossRef] [PubMed]
- WoRMS, E.B. World Register of Marine Species. Available online: https://www.marinespecies.org/ (accessed on 21 March 2022).
- Voolstra, C.R.; Quigley, K.M.; Davies, S.W.; Parkinson, J.E.; Peixoto, R.S.; Aranda, M.; Baker, A.C.; Barno, A.R.; Barshis, D.J.; Benzoni, F.; et al. Consensus guidelines for advancing coral holobiont genome and specimen voucher deposition. Front. Mar. Sci. 2021, 8, 701784. [Google Scholar] [CrossRef]
- Sigovini, M.; Keppel, E.; Tagliapietra, D. Open nomenclature in the biodiversity era. Methods Ecol. Evol. 2016, 7, 1217–1225. [Google Scholar] [CrossRef]
- Gélin, P.; Postaire, B.; Fauvelot, C.; Magalon, H. Reevaluating species number, distribution and endemism of the coral genus Pocillopora Lamarck, 1816 Using species delimitation methods and microsatellites. Mol. Phylogenet. Evol. 2017, 109, 430–446. [Google Scholar] [CrossRef]
- Schmidt-Roach, S.; Miller, K.J.; Lundgren, P.; Andreakis, N. With eyes wide open: A revision of species within and closely related to the Pocillopora damicornis species complex (Scleractinia; Pocilloporidae) using morphology and genetics. Zool. J. Linn. Soc. 2014, 170, 1–33. [Google Scholar] [CrossRef]
- Goffredo, S.; Chadwick-Furman, N. Comparative demography of mushroom corals (Scleractinia: Fungiidae) at Eilat, northern Red Sea. Mar. Biol. 2003, 142, 411–418. [Google Scholar] [CrossRef]
- Al Tawaha, M.; Benzoni, F.; Eid, E.; Abu Awali, A. Corals of Jordan: A Field Guide; The Royal Marine Conservation Society of Jordan: Amman, Jordan, 2019; ISBN 978-9957-8740-4-9. [Google Scholar]
- Apprill, A.; Hughen, K.; Mincer, T. Major similarities in the bacterial communities associated with lesioned and healthy Fungiidae corals: Healthy and lesioned Fungiid coral-bacteria. Environ. Microbiol. 2013, 15, 2063–2072. [Google Scholar] [CrossRef] [PubMed]
- Lampert, Y.; Kelman, D.; Dubinsky, Z.; Nitzan, Y.; Hill, R.T. Diversity of culturable bacteria in the mucus of the Red Sea coral Fungia scutaria. FEMS Microbiol. Ecol. 2006, 58, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Allers, E.; Niesner, C.; Wild, C.; Pernthaler, J. Microbes enriched in seawater after addition of coral mucus. Appl. Environ. Microbiol. 2008, 74, 3274–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampert, Y.; Kelman, D.; Nitzan, Y.; Dubinsky, Z.; Behar, A.; Hill, R.T. Phylogenetic diversity of bacteria associated with the mucus of red sea corals: Diversity of bacteria associated with coral mucus. FEMS Microbiol. Ecol. 2008, 64, 187–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeevi Ben Yosef, D.; Ben-Dov, E.; Kushmaro, A. Amorphus coralli gen. nov., sp. nov., a Marine bacterium isolated from coral mucus, belonging to the order Rhizobiales. Int. J. Syst. Evol. Microbiol. 2008, 58, 2704–2709. [Google Scholar] [CrossRef] [Green Version]
- Shnit-Orland, M.; Kushmaro, A. Coral mucus-associated bacteria: A possible first line of defense. FEMS Microbiol. Ecol. 2009, 67, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Ben-Dov, E.; Ben Yosef, D.Z.; Pavlov, V.; Kushmaro, A. Corynebacterium maris sp. nov., a marine bacterium isolated from the mucus of the coral Fungia granulosa. Int. J. Syst. Evol. Microbiol. 2009, 59, 2458–2463. [Google Scholar] [CrossRef] [PubMed]
- Al-Zereini, W.; Fotso Fondja Yao, C.B.; Laatsch, H.; Anke, H. Aqabamycins A-G: Novel nitro maleimides from a marine Vibrio species. I. Taxonomy, fermentation, isolation and biological activities. J. Antibiot. 2010, 63, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Shnit-Orland, M.; Sivan, A.; Kushmaro, A. Shewanella corallii sp. nov., a marine bacterium isolated from a Red Sea coral. Int. J. Syst. Evol. Microbiol. 2010, 60, 2293–2297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meron, D.; Atias, E.; Iasur Kruh, L.; Elifantz, H.; Minz, D.; Fine, M.; Banin, E. The impact of reduced PH on the microbial community of the coral Acropora eurystoma. ISME J. 2011, 5, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, O.O.; Yang, J.; Bougouffa, S.; Wang, Y.; Batang, Z.; Tian, R.; Al-Suwailem, A.; Qian, P.-Y. Spatial and species variations in bacterial communities associated with corals from the Red Sea as revealed by pyrosequencing. Appl. Environ. Microbiol. 2012, 78, 7173–7184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shnit-Orland, M.; Sivan, A.; Kushmaro, A. Antibacterial activity of Pseudoalteromonas in the coral holobiont. Microb. Ecol. 2012, 64, 851–859. [Google Scholar] [CrossRef] [PubMed]
- El-Bondkly, A.-M.; El-Gendy, M.M.A.A.; Wiese, J.; Imhoff, J.F. Phylogenetic diversity and antimicrobial activities of culturable endophytic Actinobacteria isolated from different egyptian marine sponges and soft corals. Aust. J. Basic Appl. Sci. 2012, 6, 25–33. [Google Scholar]
- Paramasivam, N.; Ben-Dov, E.; Arotsker, L.; Kramarsky-Winter, E.; Zvuloni, A.; Loya, Y.; Kushmaro, A. Bacterial consortium of Millepora dichotoma exhibiting unusual multifocal lesion event in the Gulf of Eilat, Red Sea. Microb. Ecol. 2013, 65, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Jessen, C.; Villa Lizcano, J.F.; Bayer, T.; Roder, C.; Aranda, M.; Wild, C.; Voolstra, C.R. In-situ effects of eutrophication and overfishing on physiology and bacterial diversity of the Red Sea coral Acropora hemprichii. PLoS ONE 2013, 8, e62091. [Google Scholar] [CrossRef]
- Bayer, T.; Neave, M.J.; Alsheikh-Hussain, A.; Aranda, M.; Yum, L.K.; Mincer, T.; Hughen, K.; Apprill, A.; Voolstra, C.R. The microbiome of the Red Sea coral Stylophora pistillata is dominated by tissue-associated Endozoicomonas bacteria. Appl. Environ. Microbiol. 2013, 79, 4759–4762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ElAhwany, A.M.D.; Ghozlan, H.A.; ElSharif, H.A.; Sabry, S.A. Phylogenetic diversity and antimicrobial activity of marine bacteria associated with the soft coral Sarcophyton glaucum. J. Basic Microbiol. 2013, 55, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Roder, C.; Bayer, T.; Aranda, M.; Kruse, M.; Voolstra, C.R. Microbiome structure of the Fungid coral Ctenactis echinata aligns with environmental differences. Mol. Ecol. 2015, 24, 3501–3511. [Google Scholar] [CrossRef] [PubMed]
- Röthig, T.; Ochsenkühn, M.A.; Roik, A.; van der Merwe, R.; Voolstra, C.R. Long-term salinity tolerance is accompanied by major restructuring of the coral bacterial microbiome. Mol. Ecol. 2016, 25, 1308–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, M.; Roik, A.; Porter, A.; Zubier, K.; Mudarris, M.S.; Ormond, R.; Voolstra, C.R. Coral Microbial Community dynamics in response to anthropogenic impacts near a major city in the central Red Sea. Mar. Pollut. Bull. 2016, 105, 629–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadaidi, G.; Röthig, T.; Yum, L.K.; Ziegler, M.; Arif, C.; Roder, C.; Burt, J.; Voolstra, C.R. Stable mucus-associated bacterial communities in bleached and healthy corals of Porites lobata from the Arabian Seas. Sci. Rep. 2017, 7, 45362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röthig, T.; Roik, A.; Yum, L.K.; Voolstra, C.R. Distinct bacterial microbiomes associate with the deep-sea coral Eguchipsammia fistula from the Red Sea and from aquaria settings. Front. Mar. Sci. 2017, 4, 259. [Google Scholar] [CrossRef] [Green Version]
- Röthig, T.; Yum, L.K.; Kremb, S.G.; Roik, A.; Voolstra, C.R. Microbial community composition of deep-sea corals from the Red Sea provides insight into functional adaption to a unique environment. Sci. Rep. 2017, 7, 44714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogoreutz, C.; Rädecker, N.; Cárdenas, A.; Gärdes, A.; Voolstra, C.R.; Wild, C. Sugar enrichment provides evidence for a role of nitrogen fixation in coral bleaching. Glob. Change Biol. 2017, 23, 3838–3848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Samak, M.; Solyman, S.M.; Hanora, A. Antimicrobial activity of bacteria isolated from Red Sea marine invertebrates. Biotechnol. Rep. 2018, 19, e00275. [Google Scholar] [CrossRef] [PubMed]
- Helal, H.S.; Hanora, A.; Khattab, R.A.; Hamouda, H.; Zedan, H. Mining of Egypt’s Red Sea invertebrates for potential bioactive producers. Biotechnol. Lett. 2018, 40, 1519–1530. [Google Scholar] [CrossRef]
- Freire, I.; Gutner-Hoch, E.; Muras, A.; Benayahu, Y.; Otero, A. The effect of bacteria on planula-larvae settlement and metamorphosis in the octocoral Rhytisma fulvum fulvum. PLoS ONE 2019, 14, e0223214. [Google Scholar] [CrossRef]
- Hussien, E.; Juhmani, A.-S.; AlMasri, R.; Al-Horani, F.; Al-Saghir, M. Metagenomic analysis of microbial community associated with coral mucus from the Gulf of Aqaba. Heliyon 2019, 5, e02876. [Google Scholar] [CrossRef] [PubMed]
- Cleary, D.F.R.; Polónia, A.R.M.; Reijnen, B.T.; Berumen, M.L.; de Voogd, N.J. Prokaryote communities inhabiting endemic and newly discovered sponges and octocorals from the red sea. Microb. Ecol. 2020, 80, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Gamaleldin, N.M.; Bakeer, W.; Sayed, A.M.; Shamikh, Y.I.; El-Gendy, A.O.; Hassan, H.M.; Horn, H.; Abdelmohsen, U.R.; Hozzein, W.N. Exploration of chemical diversity and antitrypanosomal activity of some red sea-derived actinomycetes using the OSMAC approach supported by lc-ms-based metabolomics and molecular modelling. Antibiotics 2020, 9, 629. [Google Scholar] [CrossRef] [PubMed]
- Meron, D.; Maor-Landaw, K.; Eyal, G.; Elifantz, H.; Banin, E.; Loya, Y.; Levy, O. The complexity of the holobiont in the red sea coral Euphyllia paradivisa under heat stress. Microorganisms 2020, 8, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngugi, D.K.; Ziegler, M.; Duarte, C.M.; Voolstra, C.R. Genomic blueprint of glycine betaine metabolism in coral metaorganisms and their contribution to reef nitrogen budgets. iScience 2020, 23, 101120. [Google Scholar] [CrossRef] [PubMed]
- Voolstra, C.R.; Valenzuela, J.J.; Turkarslan, S.; Cárdenas, A.; Hume, B.C.C.; Perna, G.; Buitrago-López, C.; Rowe, K.; Orellana, M.V.; Baliga, N.S.; et al. Contrasting heat stress response patterns of coral holobionts across the Red Sea suggest distinct mechanisms of thermal tolerance. Mol. Ecol. 2021, 30, 4466–4480. [Google Scholar] [CrossRef]
- Savary, R.; Barshis, D.; Voolstra, C.; Cardenas, A.; Evensen, N.; Banc-Prandi, G.; Fine, M.; Meibom, A. Fast and pervasive transcriptomic resilience and acclimation of extremely heat-tolerant coral holobionts from the northern Red Sea. Proc. Natl. Acad. Sci. USA 2021, 118, e2023298118. [Google Scholar] [CrossRef]
- Ben-Haim, Y.; Thompson, F.L.; Thompson, C.C.; Cnockaert, M.C.; Hoste, B.; Swings, J.; Rosenberg, E. Vibrio coralliilyticus sp. nov., a temperature-dependent pathogen of the coral Pocillopora damicornis. Int. J. Syst. Evol. Microbiol. 2003, 53, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Barash, Y.; Sulam, R.; Loya, Y.; Rosenberg, E. Bacterial strain BA-3 and a filterable factor cause a White Plague-like Disease in corals from the Eilat coral reef. Aquat. Microb. Ecol. 2005, 40, 183–189. [Google Scholar] [CrossRef]
- Thompson, F.L.; Barash, Y.; Sawabe, T.; Sharon, G.; Swings, J.; Rosenberg, E.Y. Thalassomonas loyana sp. nov., a causative agent of the White Plague-like Disease of corals on the Eilat coral reef. Int. J. Syst. Evol. Microbiol. 2006, 56, 365–368. [Google Scholar] [CrossRef] [Green Version]
- Barneah, O.; Ben-Dov, E.; Kramarsky-Winter, E.; Kushmaro, A. Characterization of black band disease in red sea stony corals. Environ. Microbiol. 2007, 9, 1995–2006. [Google Scholar] [CrossRef]
- Rasoulouniriana, D.; Siboni, N.; Ben-Dov, E.; Kramarsky-Winter, E.; Loya, Y.; Kushmaro, A. Pseudoscillatoria coralii gen. nov., sp. nov., a Cyanobacterium associated with coral Black Band Disease (BBD). Dis. Aquat. Organ. 2009, 87, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Arotsker, L.; Siboni, N.; Ben-Dov, E.; Kramarsky-Winter, E.; Loya, Y.; Kushmaro, A. Vibrio sp. as a Potentially important member of the Black Band Disease (BBD) consortium in Favia sp. Corals. FEMS Microbiol. Ecol. 2009, 70, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paramasivam, N.; Ben-Dov, E.; Arotsker, L.; Kushmaro, A. Eilatimonas milleporae gen. nov., sp. nov., a Marine bacterium isolated from the hydrocoral Millepora dichotoma. Int. J. Syst. Evol. Microbiol. 2013, 63, 1880–1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arotsker, L.; Kramarsky-Winter, E.; Ben-Dov, E.; Siboni, N.; Kushmaro, A. Changes in the bacterial community associated with Black Band Disease in a Red Sea coral, Favia sp., in relation to disease phases. Dis. Aquat. Organ. 2015, 116, 47–58. [Google Scholar] [CrossRef]
- Hadaidi, G.; Ziegler, M.; Shore-Maggio, A.; Jensen, T.; Aeby, G.; Voolstra, C.R. Ecological and molecular characterization of a coral Black Band Disease outbreak in the Red Sea during a bleaching event. PeerJ 2018, 6, e5169. [Google Scholar] [CrossRef] [Green Version]
- Roik, A.; Röthig, T.; Roder, C.; Ziegler, M.; Kremb, S.G.; Voolstra, C.R. Year-long monitoring of physico-chemical and biological variables provide a comparative baseline of coral reef functioning in the central Red Sea. PLoS ONE 2016, 11, e0163939. [Google Scholar] [CrossRef] [Green Version]
- Becker, C.; Hughen, K.; Mincer, T.J.; Ossolinski, J.; Weber, L.; Apprill, A. Impact of prawn farming effluent on coral reef water nutrients and microorganisms. Aquac. Environ. Interact. 2017, 9, 331–346. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas, A.; Neave, M.J.; Haroon, M.F.; Pogoreutz, C.; Rädecker, N.; Wild, C.; Gärdes, A.; Voolstra, C.R. excess labile carbon promotes the expression of virulence factors in coral reef bacterioplankton. ISME J. 2018, 12, 59–76. [Google Scholar] [CrossRef] [Green Version]
- Pearman, J.K.; Aylagas, E.; Voolstra, C.R.; Anlauf, H.; Villalobos, R.; Carvalho, S. Disentangling the complex microbial community of coral reefs using standardized Autonomous Reef Monitoring Structures (ARMS). Mol. Ecol. 2019, 28, 3496–3507. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, R.S.; Sweet, M.; Bourne, D.G. Customized medicine for corals. Front. Mar. Sci. 2019, 6, 686. [Google Scholar] [CrossRef]
- Peixoto, R.S.; Sweet, M.; Villela, H.D.M.; Cardoso, P.; Thomas, T.; Voolstra, C.R.; Høj, L.; Bourne, D.G. Coral Probiotics: Premise, promise, prospects. Annu. Rev. Anim. Biosci. 2021, 9, 265–288. [Google Scholar] [CrossRef]
- Voolstra, C.R.; Suggett, D.J.; Peixoto, R.S.; Parkinson, J.E.; Quigley, K.M.; Silveira, C.B.; Sweet, M.; Muller, E.M.; Barshis, D.J.; Bourne, D.G.; et al. Extending the natural adaptive capacity of coral holobionts. Nat. Rev. Earth Environ. 2021, 2, 747–762. [Google Scholar] [CrossRef]
- Doering, T.; Wall, M.; Putchim, L.; Rattanawongwan, T.; Schroeder, R.; Hentschel, U.; Roik, A. Towards enhancing coral heat tolerance: A “Microbiome transplantation” treatment using inoculations of homogenized coral tissues. Microbiome 2021, 9, 102. [Google Scholar] [CrossRef] [PubMed]
- Morgans, C.A.; Hung, J.Y.; Bourne, D.G.; Quigley, K.M. Symbiodiniaceae probiotics for use in bleaching recovery. Restor. Ecol. 2020, 28, 282–288. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Q.; Ling, J.; Long, L.; Huang, H.; Yin, J.; Wu, M.; Tang, X.; Lin, X.; Zhang, Y.; et al. Shifting the microbiome of a coral holobiont and improving host physiology by inoculation with a potentially beneficial bacterial consortium. BMC Microbiol. 2021, 21, 130. [Google Scholar] [CrossRef]
- Sofianos, S.S.; Johns, W.E. An Oceanic General Circulation Model (OGCM) Investigation of the Red Sea circulation, 1. Exchange between the Red Sea and the Indian ocean. J. Geophys. Res. Oceans 2002, 107, 17-1–17-11. [Google Scholar] [CrossRef]
- Berumen, M.L.; Hoey, A.S.; Bass, W.H.; Bouwmeester, J.; Catania, D.; Cochran, J.E.M.; Khalil, M.T.; Miyake, S.; Mughal, M.R.; Spaet, J.L.Y.; et al. The status of coral reef ecology research in the Red Sea. Coral Reefs 2013, 32, 737–748. [Google Scholar] [CrossRef]
- Aeby, G.S.; Shore, A.; Jensen, T.; Ziegler, M.; Work, T.; Voolstra, C.R. A comparative baseline of coral disease in three regions along the Saudi Arabian Coast of the central Red Sea. PLoS ONE 2021, 16, e0246854. [Google Scholar] [CrossRef] [PubMed]
- Terraneo, T.I.; Benzoni, F.; Baird, A.H.; Arrigoni, R.; Berumen, M.L. Morphology and molecules reveal two new species of Porites (Scleractinia, Poritidae) from the Red Sea and the Gulf of Aden. Syst. Biodivers. 2019, 17, 491–508. [Google Scholar] [CrossRef]
- Devantier, L.; Turak, E.; Al-Shaikh, K.; De’ath, G. Coral communities of the central-northern Saudi Arabian Red Sea. Fauna Arab. 2000, 18, 23–66. [Google Scholar]
- Dar, M.A.; Mohamad, T.A.; El-Saman, M.I. Geomorphic and geographic distributions of reef-building and soft coral communities in the inshore zone of the Red Sea. Egypt. J. Aquat. Res. 2012, 38, 105–117. [Google Scholar] [CrossRef]
- Hegazy, M.E.F.; Mohamed, T.A.; Alhammady, M.A.; Shaheen, A.M.; Reda, E.H.; Elshamy, A.I.; Aziz, M.; Paré, P.W. Molecular architecture and biomedical leads of terpenes from Red Sea marine invertebrates. Mar. Drugs 2015, 13, 3154–3181. [Google Scholar] [CrossRef] [PubMed]
- Afifi, R.; Abdel-Nabi, I.M.; El-Shaikh, K. Antibacterial activity from soft corals of the Red Sea, Saudi Arabia. J. Taibah Univ. Sci. 2016, 10, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Farag, M.A.; Porzel, A.; Al-Hammady, M.A.; Hegazy, M.-E.F.; Meyer, A.; Mohamed, T.A.; Westphal, H.; Wessjohann, L.A. Soft corals biodiversity in the Egyptian Red Sea: A comparative MS and NMR metabolomics approach of wild and aquarium grown species. J. Proteome Res. 2016, 15, 1274–1287. [Google Scholar] [CrossRef] [PubMed]
- Raimundo, I.; Silva, S.G.; Costa, R.; Keller-Costa, T. Bioactive secondary metabolites from octocoral-associated microbes—New chances for blue growth. Mar. Drugs 2018, 16, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modolon, F.; Barno, A.; Villela, H.; Peixoto, R. Ecological and biotechnological importance of secondary metabolites produced by coral-associated bacteria. J. Appl. Microbiol. 2020, 129, 1441–1457. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.H.; Zidan, S.A.H.; Samy, M.N.; Alian, A.; Abdelmohsen, U.R.; Fouad, M.A.; Kamel, M.S.; Matsunami, K. Cytotoxicity and chemical profiling of the Red Sea soft corals Litophyton arboreum. Nat. Prod. Res. 2021, 36, 4261–4265. [Google Scholar] [CrossRef] [PubMed]
- Alian, A.A.; Abdel Shakor, A.B.; -Geba, K.M.; Osman, A.G.M. Cytotoxicity and antimicrobial activities of some soft corals inhabiting the Red Sea, Egypt. Al-Azhar Bull. Sci. 2021, 32, 1–7. [Google Scholar] [CrossRef]
- Benayahu, Y. Reproduction and developmental pathways of Red Sea Xeniidae (Octocorallia, Alcyonacea). Hydrobiologia 1991, 216, 125–130. [Google Scholar] [CrossRef]
- Reinicke, G.B. Xeniidae Des Roten Meeres:(Octocorallia, Alcyonacea); Beiträge Zur Systematik Und Ökologie; Westarp Wiss: Magdeburg, Germany, 1995. [Google Scholar]
- Benayahu, Y.; McFadden, C.S. A New genus of soft coral of the family Alcyoniidae (Cnidaria, Octocorallia) with re-description of a new combination and description of a new species. ZooKeys 2011, 84, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Benayahu, Y.; McFadden, C.S.; Shoham, E. Search for mesophotic octocorals (Cnidaria, Anthozoa) and their phylogeny: I. A new sclerite-free genus from Eilat, northern Red Sea. ZooKeys 2017, 680, 1–11. [Google Scholar] [CrossRef] [PubMed]
- van de Water, J.A.J.M.; Allemand, D.; Ferrier-Pagès, C. Host-microbe interactions in octocoral holobionts-recent advances and perspectives. Microbiome 2018, 6, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakimov, M.M.; Cappello, S.; Crisafi, E.; Tursi, A.; Savini, A.; Corselli, C.; Scarfi, S.; Giuliano, L. Phylogenetic survey of metabolically active microbial communities associated with the deep-sea coral Lophelia pertusa from the Apulian Plateau, central Mediterranean Sea. Deep Sea Res. Part Oceanogr. Res. Pap. 2006, 53, 62–75. [Google Scholar] [CrossRef]
- Neulinger, S.C.; Järnegren, J.; Ludvigsen, M.; Lochte, K.; Dullo, W.-C. Phenotype-specific bacterial communities in the cold-water coral Lophelia pertusa (Scleractinia) and their implications for the coral’s nutrition, health, and distribution. Appl. Environ. Microbiol. 2008, 74, 7272–7285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roder, C.; Berumen, M.L.; Bouwmeester, J.; Papathanassiou, E.; Al-Suwailem, A.; Voolstra, C.R. First biological measurements of deep-sea corals from the red sea. Sci. Rep. 2013, 3, 2802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meistertzheim, A.-L.; Lartaud, F.; Arnaud-Haond, S.; Kalenitchenko, D.; Bessalam, M.; Le Bris, N.; Galand, P.E. Patterns of bacteria-host associations suggest different ecological strategies between two reef building cold-water coral species. Deep Sea Res. Part Oceanogr. Res. Pap. 2016, 114, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Roik, A.; Röthig, T.; Roder, C.; Müller, P.J.; Voolstra, C.R. Captive rearing of the deep-sea coral Eguchipsammia fistula from the Red Sea demonstrates remarkable physiological plasticity. PeerJ 2015, 3, e734. [Google Scholar] [CrossRef] [Green Version]
- Olson, J.B.; Kellogg, C.A. Microbial ecology of corals, sponges, and algae in mesophotic coral environments: Mesophotic microbial ecology. FEMS Microbiol. Ecol. 2010, 73, 17–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, E.O.; Weinnig, A.M. Microbiomes and obligate symbiosis of deep-sea animals. Annu. Rev. Anim. Biosci. 2022, 10, 151–176. [Google Scholar] [CrossRef] [PubMed]
- Beydoun, Z.R. The hydrocarbon prospects of the red sea-gulf of aden: A Review. J. Pet. Geol. 1989, 12, 125–144. [Google Scholar] [CrossRef]
- Thompson, F.L.; Iida, T.; Swings, J. Biodiversity of Vibrios. Microbiol. Mol. Biol. Rev. 2004, 68, 403–431. [Google Scholar] [CrossRef] [PubMed]
- Sunagawa, S.; DeSantis, T.Z.; Piceno, Y.M.; Brodie, E.L.; DeSalvo, M.K.; Voolstra, C.R.; Weil, E.; Andersen, G.L.; Medina, M. Bacterial diversity and white plague disease-associated community changes in the caribbean Coral Montastraea faveolata. ISME J. 2009, 3, 512–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dryselius, R.; Kurokawa, K.; Iida, T. Vibrionaceae, a versatile bacterial family with evolutionarily conserved variability. Res. Microbiol. 2007, 158, 479–486. [Google Scholar] [CrossRef]
- Gregory, G.J.; Boyd, E.F. Stressed out: Bacterial response to high salinity using compatible solute biosynthesis and uptake systems, lessons from Vibrionaceae. Comput. Struct. Biotechnol. J. 2021, 19, 1014–1027. [Google Scholar] [CrossRef] [PubMed]
- Raimundo, I.; Silva, R.; Meunier, L.; Valente, S.M.; Lago-Lestón, A.; Keller-Costa, T.; Costa, R. Functional metagenomics reveals differential chitin degradation and utilization features across free-living and host-associated marine microbiomes. Microbiome 2021, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Zhenyu, X.; Shaowen, K.; Chaoqun, H.; Zhixiong, Z.; Shifeng, W.; Yongcan, Z. First characterization of bacterial pathogen, Vibrio Alginolyticus, for Porites andrewsi White Syndrome in the South China Sea. PLoS ONE 2013, 8, e75425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovanni, S.J.; Rappé, M.S. Evolution, diversity and molecular ecology of marine prokaryotes. In Microbial Ecology of the Oceans; John Wiley & Sons: Hoboken, N, USA, 2000; pp. 47–84. [Google Scholar]
- Pootakham, W.; Mhuantong, W.; Yoocha, T.; Putchim, L.; Jomchai, N.; Sonthirod, C.; Naktang, C.; Kongkachana, W.; Tangphatsornruang, S. Heat-induced shift in coral microbiome reveals several members of the Rhodobacteraceae family as indicator species for thermal stress in Porites lutea. MicrobiologyOpen 2019, 8, e935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apprill, A.; Marlow, H.Q.; Martindale, M.Q.; Rappé, M.S. The onset of microbial associations in the coral Pocillopora meandrina. ISME J. 2009, 3, 685–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, N.; Motone, K.; Takagi, T.; Aburaya, S.; Watanabe, S.; Aoki, W.; Ueda, M. Ruegeria sp. Strains isolated from the reef-building coral Galaxea fascicularis inhibit growth of the temperature-dependent pathogen Vibrio coralliilyticus. Mar. Biotechnol. 2019, 21, 1–8. [Google Scholar] [CrossRef]
- Luo, D.; Wang, X.; Feng, X.; Tian, M.; Wang, S.; Tang, S.-L.; Ang, P.; Yan, A.; Luo, H. Population differentiation of Rhodobacteraceae along with coral compartments. ISME J. 2021, 15, 3286–3302. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.; Warnecke, F.; Amann, R.; Pernthaler, J. High local and global diversity of Flavobacteria in marine plankton. Environ. Microbiol. 2007, 9, 1253–1266. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, X.-Y.; Wen, X.-R.; Shi, M.; Chen, X.-L.; Su, H.-N. Arcticiflavibacter luteus gen. nov., sp. nov., a member of the family Flavobacteriaceae isolated from intertidal sand. Int. J. Syst. Evol. Microbiol. 2016, 66, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Nedashkovskaya, O.I.; Kim, S.B.; Lysenko, A.M.; Frolova, G.M.; Mikhailov, V.V.; Bae, K.S. Bizionia paragorgiae gen. nov., sp. nov., a novel member of the family Flavobacteriaceae isolated from the soft coral Paragorgia arborea. Int. J. Syst. Evol. Microbiol. 2005, 55, 375–378. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.-M.; Giovannoni, S.J.; Lee, K.; Ferriera, S.; Johnson, J.; Cho, J.-C. Complete genome sequence of Robiginitalea biformata HTCC2501. J. Bacteriol. 2009, 191, 7144–7145. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.-X.; Du, Z.-J.; Chen, G.-J. Seonamhaeicola algicola sp. nov., a complex-polysaccharide-degrading bacterium isolated from Gracilaria blodgettii, and emended description of the genus Seonamhaeicola. Int. J. Syst. Evol. Microbiol. 2016, 66, 2064–2068. [Google Scholar] [CrossRef] [Green Version]
- Yin, R.; Yi, Y.-J.; Chen, Z.; Chen, G.-J.; Zhou, Y.-X.; Du, Z.-J. Flavihalobacter algicola gen. nov. sp. nov., a member of the family Flavobacteriaceae with alginate-degradation activity, isolated from marine alga Saccharina japonica. Int. J. Syst. Evol. Microbiol. 2021, 71, 004701. [Google Scholar] [CrossRef]
- Raina, J.-B.; Dinsdale, E.A.; Willis, B.L.; Bourne, D.G. Do the organic sulfur compounds DMSP and DMS drive coral microbial associations? Trends Microbiol. 2010, 18, 101–108. [Google Scholar] [CrossRef]
- Raina, J.-B.; Tapiolas, D.M.; Forêt, S.; Lutz, A.; Abrego, D.; Ceh, J.; Seneca, F.O.; Clode, P.L.; Bourne, D.G.; Willis, B.L.; et al. DMSP biosynthesis by an animal and its role in coral thermal stress response. Nature 2013, 502, 677–680. [Google Scholar] [CrossRef] [Green Version]
- Howard, E.C.; Sun, S.; Reisch, C.R.; del Valle, D.A.; Bürgmann, H.; Kiene, R.P.; Moran, M.A. Changes in dimethylsulfoniopropionate demethylase gene assemblages in response to an induced phytoplankton bloom. Appl. Environ. Microbiol. 2011, 77, 524–531. [Google Scholar] [CrossRef] [Green Version]
- Raina, J.-B.; Tapiolas, D.; Motti, C.A.; Foret, S.; Seemann, T.; Tebben, J.; Willis, B.L.; Bourne, D.G. Isolation of an antimicrobial compound produced by bacteria associated with reef-building corals. PeerJ 2016, 4, e2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neave, M.J.; Apprill, A.; Ferrier-Pagès, C.; Voolstra, C.R. Diversity and function of prevalent symbiotic marine bacteria in the genus Endozoicomonas. Appl. Microbiol. Biotechnol. 2016, 100, 8315–8324. [Google Scholar] [CrossRef] [PubMed]
- Tandon, K.; Lu, C.-Y.; Chiang, P.-W.; Wada, N.; Yang, S.-H.; Chan, Y.-F.; Chen, P.-Y.; Chang, H.-Y.; Chiou, Y.-J.; Chou, M.-S.; et al. Comparative genomics: Dominant coral-bacterium Endozoicomonas acroporae metabolizes dimethylsulfoniopropionate (DMSP). ISME J. 2020, 14, 1290–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantos, O.; Bongaerts, P.; Dennis, P.G.; Tyson, G.W.; Hoegh-Guldberg, O. Habitat-specific environmental conditions primarily control the microbiomes of the coral Seriatopora hystrix. ISME J. 2015, 9, 1916–1927. [Google Scholar] [CrossRef] [PubMed]
- Lesser, M.P.; Mazel, C.H.; Gorbunov, M.Y.; Falkowski, P.G. Discovery of symbiotic nitrogen-fixing cyanobacteria in corals. Science 2004, 305, 997–1000. [Google Scholar] [CrossRef] [PubMed]
- Ceh, J.; Kilburn, M.R.; Cliff, J.B.; Raina, J.-B.; van Keulen, M.; Bourne, D.G. Nutrient cycling in early coral life stages: Pocillopora damicornis larvae provide their algal symbiont (Symbiodinium) with nitrogen acquired from bacterial associates. Ecol. Evol. 2013, 3, 2393–2400. [Google Scholar] [CrossRef]
- Brooks, A.W.; Kohl, K.D.; Brucker, R.M.; van Opstal, E.J.; Bordenstein, S.R. Phylosymbiosis: Relationships and functional effects of microbial communities across host evolutionary history. PLoS Biol. 2016, 14, e2000225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Rivière, M.; Garrabou, J.; Bally, M. Evidence for host specificity among dominant bacterial symbionts in temperate gorgonian corals. Coral Reefs 2015, 34, 1087–1098. [Google Scholar] [CrossRef]
- Cao, Y.; Dietrich, C.H. Phylogenomics of flavobacterial insect nutritional endosymbionts with implications for Auchenorrhyncha Phylogeny. Cladistics 2022, 38, 38–58. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, M.; Kurisu, F.; Kasuga, I.; Furumai, H. Time-resolved dna stable isotope probing links Desulfobacterales and Coriobacteriaceae-related bacteria to anaerobic degradation of benzene under methanogenic conditions. Microbes Environ. 2014, 29, 191–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhang, H.; Wang, X.; Yu, X.; Hu, C.; Zhang, X. The family Coriobacteriaceae is a potential contributor to the beneficial effects of roux-en-y gastric bypass on type 2 diabetes. Surg. Obes. Relat. Dis. 2018, 14, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Sadaiappan, B.; PrasannaKumar, C.; Nambiar, V.U.; Subramanian, M.; Gauns, M.U. Meta-analysis cum machine learning approaches address the structure and biogeochemical potential of marine copepod associated bacteriobiomes. Sci. Rep. 2021, 11, 3312. [Google Scholar] [CrossRef]
- Reed, M.B. Analysis of Microbial Association Networks from the Mucus of Corals and Whale Sharks. Master’s Thesis, San Diego State University, San Diego, CA, USA, 2020. [Google Scholar]
- Hördt, A.; López, M.G.; Meier-Kolthoff, J.P.; Schleuning, M.; Weinhold, L.-M.; Tindall, B.J.; Gronow, S.; Kyrpides, N.C.; Woyke, T.; Göker, M. Analysis of 1000+ type-strain genomes substantially improves taxonomic classification of Alphaproteobacteria. Front. Microbiol. 2020, 11, 468. [Google Scholar] [PubMed]
- Siewert, C.; Hess, W.R.; Duduk, B.; Huettel, B.; Reinhardt, R.; Büttner, C.; Kube, M. Complete genome determination and analysis of Acholeplasma oculi strain 19l, highlighting the loss of basic genetic features in the Acholeplasmataceae. BMC Genomics 2014, 15, 931. [Google Scholar] [CrossRef] [Green Version]
- Boyle, P.J.; Maki, J.S.; Mitchell, R. Mollicute identified in novel association with aquatic invertebrate. Curr. Microbiol. 1987, 15, 85–89. [Google Scholar] [CrossRef]
- Pontefract, A.; Zhu, T.F.; Walker, V.K.; Hepburn, H.; Lui, C.; Zuber, M.T.; Ruvkun, G.; Carr, C.E. Microbial diversity in a hypersaline sulfate lake: A terrestrial analog of ancient Mars. Front. Microbiol. 2017, 8, 1819. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.-X.; Pang, Y.-L.; Wang, Y.-C.; Li, J.-L.; Qin, S. Distribution of bacterial communities along the spatial and environmental gradients from Bohai Sea to northern Yellow Sea. PeerJ 2018, 6, e4272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misson, G.; Mainardis, M.; Marroni, F.; Peressotti, A.; Goi, D. Environmental methane emissions from seagrass wrack and evaluation of salinity effect on microbial community composition. J. Clean. Prod. 2021, 285, 125426. [Google Scholar] [CrossRef]
- Dworkin, M.; Falkow, S.; Rosenberg, E.; Schleifer, K.-H.; Stackebrandt, E. (Eds.) The Prokaryotes; Springer: New York, NY, USA, 2006; ISBN 978-0-387-25493-7. [Google Scholar]
- Chang, X.; Liu, W.; Zhang, X.-H. Salinactinospora qingdaonensis gen. nov., sp. nov., a halophilic Actinomycete isolated from a salt pond. Int. J. Syst. Evol. Microbiol. 2012, 62, 954–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, S.-K.; Tian, X.-P.; Zhi, X.-Y.; Cai, M.; Wu, J.-Y.; Yang, L.-L.; Xu, L.-H.; Li, W.-J. Haloactinospora alba gen. nov., sp. nov., a halophilic filamentous Actinomycete of the family Nocardiopsaceae. Int. J. Syst. Evol. Microbiol. 2008, 58, 2075–2080. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.-P.; Tang, S.-K.; Dong, J.-D.; Zhang, Y.-Q.; Xu, L.-H.; Zhang, S.; Li, W.-J. Marinactinospora thermotolerans gen. nov., sp. nov., a marine Actinomycete isolated from a sediment in the northern south China Sea. Int. J. Syst. Evol. Microbiol. 2009, 59, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Han, C.; Lee, T.G.; Chin, J.; Choi, H.; Lee, W.; Paik, M.J.; Won, D.H.; Jeong, G.; Ko, J.; et al. Marinopyrones A–D, α-pyrones from marine-derived Actinomycetes of the family Nocardiopsaceae. Tetrahedron Lett. 2016, 57, 1997–2000. [Google Scholar] [CrossRef]
- Jose, P.A.; Jha, B. Intertidal marine sediment harbours Actinobacteria with promising bioactive and biosynthetic potential. Sci. Rep. 2017, 7, 10041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, Z.Y.; Tan, G.Y.A. Selective isolation and characterization of novel members of the family Nocardiopsaceae and other Actinobacteria from a marine sediment of Tioman island. Antonie Van Leeuwenhoek 2018, 111, 727–742. [Google Scholar] [CrossRef]
- Goffredi, S.K. Indigenous ectosymbiotic bacteria associated with diverse hydrothermal vent invertebrates. Environ. Microbiol. Rep. 2010, 2, 479–488. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef]
- Hugenholtz, P.; Goebel, B.M.; Pace, N.R. Impact of culture-independent studies on the emerging phylogenetic view of bacterial diversity. J. Bacteriol. 1998, 180, 4765–4774. [Google Scholar] [CrossRef] [Green Version]
- Molina-Menor, E.; Gimeno-Valero, H.; Pascual, J.; Peretó, J.; Porcar, M. High culturable bacterial diversity from a european desert: The Tabernas desert. Front. Microbiol. 2021, 11, 583120. [Google Scholar] [CrossRef]
- Lagier, J.-C.; Khelaifia, S.; Alou, M.T.; Ndongo, S.; Dione, N.; Hugon, P.; Caputo, A.; Cadoret, F.; Traore, S.I.; Seck, E.H.; et al. Culture of previously uncultured members of the human gut microbiota by culturomics. Nat. Microbiol. 2016, 1, 16203. [Google Scholar] [CrossRef] [PubMed]
- Berdy, B.; Spoering, A.L.; Ling, L.L.; Epstein, S.S. In situ cultivation of previously uncultivable microorganisms using the Ichip. Nat. Protoc. 2017, 12, 2232–2242. [Google Scholar] [CrossRef] [PubMed]
- Martiny, A.C. High proportions of bacteria are culturable across major biomes. ISME J. 2019, 13, 2125–2128. [Google Scholar] [CrossRef] [PubMed]
- Bernard, G.; Pathmanathan, J.S.; Lannes, R.; Lopez, P.; Bapteste, E. Microbial Dark Matter Investigations: How microbial studies transform biological knowledge and empirically sketch a logic of scientific discovery. Genome Biol. Evol. 2018, 10, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, R. The Red Sea Research Center Microbial Biovault. 2022. Available online: https://marinemicrobiomeslab.kaust.edu.sa/research/detail/the-red-sea-research-center-microbial-biovault (accessed on 10 November 2022).
- Peixoto, R. The Beneficial Microbes for Marine Organisms (BMMO) Network. 2017. Available online: http://bmmo.microbe.net/ (accessed on 10 November 2022).
- Neave, M.J.; Apprill, A.; Aeby, G.; Miyake, S.; Voolstra, C.R. Microbial communities of Red Sea coral reefs. In Coral reefs of the Red Sea; Voolstra, C.R., Berumen, M.L., Eds.; Coral reefs of the world; Springer International Publishing: Cham, Switzerland, 2019; pp. 53–68. ISBN 978-3-030-05802-9. [Google Scholar]
- Sawall, Y.; Al-Sofyani, A. Biology of Red Sea corals: Metabolism, reproduction, acclimatization, and adaptation. In The Red Sea: The Formation, Morphology, Oceanography and Environment of a Young Ocean Basin; Rasul, N.M.A., Stewart, I.C.F., Eds.; Springer Earth System Sciences; Springer: Berlin/Heidelberg, Germany, 2015; pp. 487–509. ISBN 978-3-662-45201-1. [Google Scholar]
- Knowlton, N. Local management matters for coral reefs. Science 2021, 372, 908–909. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Corals | ||||||
|---|---|---|---|---|---|---|
| Year | Reference | Red Sea Region | Host | Accession Number | Data Deposition | Microbial Identification Approach |
| 2006 | [50] | N | Lobactis scutaria | DQ107384-DQ107405 | NCBI | CDT |
| 2007 | [39] | N | Pleuractis granulosa | DQ117312-DQ117435 | NCBI | CIT |
| 2008 | [51] | N | Fungia sp. | AM941168—AM941189 | NCBI | CIT |
| 2008 | [52] | N | Lobactis scutaria and Platygyra lamellina | EF466016–EF466058; EF576992–EF577027 | NCBI | CIT |
| 2008 | [53] | N | Pleuractis granulosa | DQ097300 | NCBI | CDT |
| 2009 | [54] | N | Platygyra sp., Porites sp., Pleuractis granulosa, Favia sp., Stylophora sp., Pocillopora sp., Rhytisma fulvum fulvum and Xenia sp. | FJ041063—FJ041108 | NCBI | CDT |
| 2009 | [55] | N | Pleuractis granulosa | DSM 45190 | NCBI | CDT |
| 2010 | [56] | N | Sinularia polydactyla | - | - | CDT |
| 2010 | [57] | N | Favia sp. | FJ041083 | NCBI | CDT |
| 2011 | [58] | N | Acropora eurystoma | GU319121–GU319777 | NCBI | CDT/CIT |
| 2012 | [59] | C | Pocillopora verrucosa, Astreopora myriophthalma, Stylophora pistillata, Sarcophyton sp. 1 and S. sp. 2 | SRA012656 | NCBI | CIT |
| 2012 | [60] | N | Platygyra sp., Porites sp., Stylophora sp., and Pocillopora sp. | FJ041071; FJ041068; FJ041064; FJ041069; FJ041065 | NCBI | CDT |
| 2012 | [61] | N | Dendronephthya sinaiensis, Dendronephthya hemprichi, Xenia sp., Lobularia sp., and Rhytisma fulvum | JF292923—JF292930 | NCBI | CDT |
| 2013 | [49] | N and S | Ctenactis cf crassa and Herpolitha limax | SAMN01939168–SAMN01939191 | NCBI | CIT |
| 2013 | [62] | N | Millepora dichotoma | HQ288552- HQ288737 | NCBI | CDT |
| 2013 | [63] | C | Acropora hemprichii | SRA062645 | NCBI | CIT |
| 2013 | [64] | S | Stylophora pistillata, Acropora humilis and Acropora damicornis | PRJNA189184 and KC668414 to KC669277 | NCBI | CIT |
| 2013 | [65] | N | Sarcophyton glaucum | JQ929053-JQ929072 | NCBI | CDT |
| 2015 | [66] | C | Ctenactis echinata | PRJNA277291 | NCBI | CIT |
| 2016 | [67] | C | Pleuractis granulosa | PRJNA282461 | NCBI | CIT |
| 2016 | [68] | C | Acropora hemprichii and Pocillopora verrucosa | PRJNA287432 | NCBI | CIT |
| 2017 | [36] | C | Stylophora pistillata, Pocillopora verrucosa, and Acropora humilis | PRJNA323666 | NCBI | CIT |
| 2017 | [37] | C | Stylophora pistillata and Pocillopora verrucosa | PRJNA280923 | NCBI | CIT |
| 2017 | [69] | C | Porites lobata | PRJNA352338 | NCBI | CIT |
| 2017 | [70] | C | Eguchipsammia fistula | PRJNA354830 | NCBI | CIT |
| 2017 | [71] | N and C | Dendrophyllia sp., Eguchipsammia fistula, and Rhizotrochus typus | PRJNA354830 | NCBI | CIT |
| 2017 | [72] | C | Pocillopora verrucosa | PRJNA335276 | NCBI | CIT |
| 2018 | [73] | N | 2 soft corals and 4 hard corals (coral ID not provided in the paper) | MH040868, MH040869, MH040870, MH040871, MH045063, MH045064, MH045263, MH045264, MH046783 and MH046784 | NCBI | CDT |
| 2018 | [25] | C | Pocillopora verrucosa | PRJNA394597 | NCBI | CIT |
| 2018 | [74] | N | Heteroxenia fuscescens, Litophyton arboerum, Sarcophyton trocheliophorum, Sinularia sp., Sacrophyton acutum and Lobophytum pauciflorum | MG757677.1 MG757675.1 MG757672.1 MG757678.1 | NCBI | CDT |
| 2019 | [75] | N | Rhytisma fulvum fulvum | - | - | CDT |
| 2019 | [18] | C | Acropora hemprichii and Pocillopora verrucosa | PRJNA491299 | NCBI | CIT |
| 2019 | [76] | N | Stylophora pistillata | - | - | CIT |
| 2020 | [21] | N | Porites nodifera, Dipsastraea favus, Pocillopora cf damicornis, Seriatopora hystrix, Xenia umbellata, and Sarcophyton trocheliophorum. | PRJNA509355 | NCBI | CIT |
| 2020 | [77] | C | Melithaea rubrinodis and Siphonogorgia sp. | SRP115467 | NCBI | CIT |
| 2020 | [78] | N | Sarcophyton glaucum | - | - | CDT |
| 2020 | [79] | N | Euphyllia paradivisa | SRP133996 | NCBI | CIT |
| 2020 | [80] | C | Acanthastrea echinata, Diploastrea heliopora, Fungia sp., Galaxea fascicularis, Mycedium elephantotus, Pavona varians, Plerogyra sinuosa, Pocillopora verrucosa, Porites lutea, Stylophora pistillata, Millepora platyphylla, Acropora cytherea, Pachyseris speciosa and Xenia sp. | PRJNA437202 | NCBI | CIT |
| 2021 | [81] | N and C | Stylophora pistillata | PRJNA681108 | NCBI | CIT |
| 2021 | [82] | N | Styllophora pistillata | PRJNA674053 | NCBI | CIT |
| 2021 | [16] | N and C | Acropora hemprichii, Acropora humilis, Acropora cytherea (Cardénas and Voolstra, 2021) | GS0145871; PRJNA343499 | NCBI | CDT/CIT |
| Diseased Corals | ||||||
| 2003 | [83] | N | Pocillopora cf damicornis | - | - | CDT |
| 2005 | [84] | N | Goniastrea sp. and Dipsastraea favus | AY643537 | EMBL | CDT |
| 2006 | [85] | N | Dipsastraea favus | CBMAI 722 | NCBI | CDT |
| 2007 | [86] | N | Favites sp. and Favia sp. | EF089403–EF089533, EF433087–EF433174, EF089519; EF089534–EF089544 | NCBI | CIT |
| 2009 | [87] | N | Favia sp. | FJ210722 | NCBI | CDT |
| 2009 | [88] | N | Favia sp. | GQ215061–GQ215096; GQ215097–GQ215227 | NCBI | CDT |
| 2013 | [89] | N | Millepora dichotoma | HQ288781 | NCBI | CDT |
| 2015 | [90] | N | Favia sp. | 4541470.3–4541481.3. | MG-RAST | CIT |
| 2018 | [91] | C | Astreopora sp., Coelastrea sp., Dipsastraea sp., Goniopora sp., Montipora sp., Pavona sp., Platygyra sp., Psammocora sp., and Gardineroseris sp. | PRJNA436216 | NCBI | CIT |
| Seawater and Microbial Biofilms Associated with Red Sea Coral Reefs | ||||||
| 2016 | [92] | C | Seawater and microbiome biofilm in Terracotta tiles | PRJNA306204 | NCBI | CIT |
| 2017 | [93] | C | Seawater from coral reefs | PRJNA357506 | NCBI | CIT |
| 2018 | [94] | C | Seawater from coral reefs | PRJNA352340 | NCBI | CIT |
| 2019 | [95] | N, C and S | Seawater and microbiome biofilm in Autonomous Reef Monitoring Structures (ARMS) | PRJNA479721 | NCBI | CIT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgadillo-Ordoñez, N.; Raimundo, I.; Barno, A.R.; Osman, E.O.; Villela, H.; Bennett-Smith, M.; Voolstra, C.R.; Benzoni, F.; Peixoto, R.S. Red Sea Atlas of Coral-Associated Bacteria Highlights Common Microbiome Members and Their Distribution across Environmental Gradients—A Systematic Review. Microorganisms 2022, 10, 2340. https://doi.org/10.3390/microorganisms10122340
Delgadillo-Ordoñez N, Raimundo I, Barno AR, Osman EO, Villela H, Bennett-Smith M, Voolstra CR, Benzoni F, Peixoto RS. Red Sea Atlas of Coral-Associated Bacteria Highlights Common Microbiome Members and Their Distribution across Environmental Gradients—A Systematic Review. Microorganisms. 2022; 10(12):2340. https://doi.org/10.3390/microorganisms10122340
Chicago/Turabian StyleDelgadillo-Ordoñez, Nathalia, Inês Raimundo, Adam R. Barno, Eslam O. Osman, Helena Villela, Morgan Bennett-Smith, Christian R. Voolstra, Francesca Benzoni, and Raquel S. Peixoto. 2022. "Red Sea Atlas of Coral-Associated Bacteria Highlights Common Microbiome Members and Their Distribution across Environmental Gradients—A Systematic Review" Microorganisms 10, no. 12: 2340. https://doi.org/10.3390/microorganisms10122340