Lymphocystis Disease Virus Infection in Clownfish Amphiprion ocellaris and Amphiprion clarkii in Taiwan

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Fish Samples
2.2. Pathology
2.3. Electron Microscopy
2.4. PCR Detection of MCP Genes
2.5. Sequence Analysis
3. Results
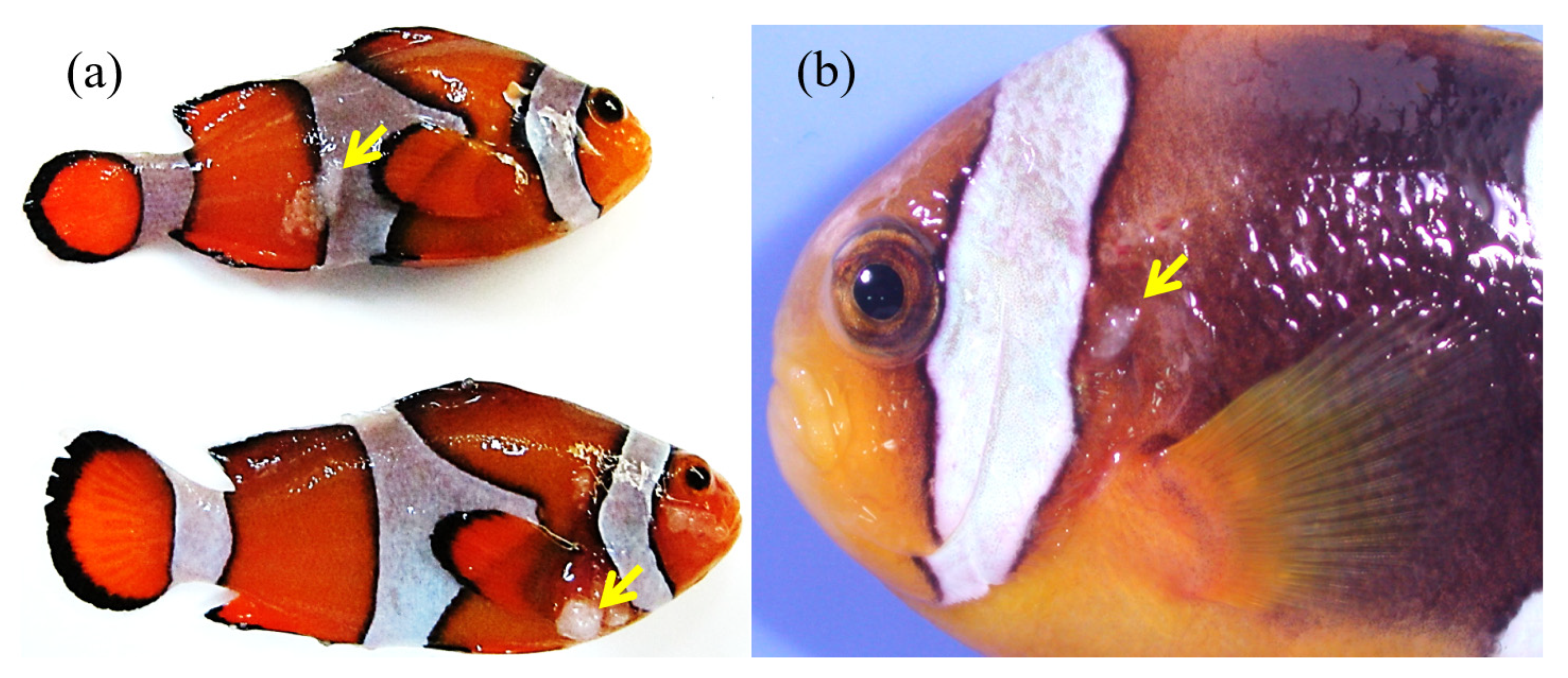
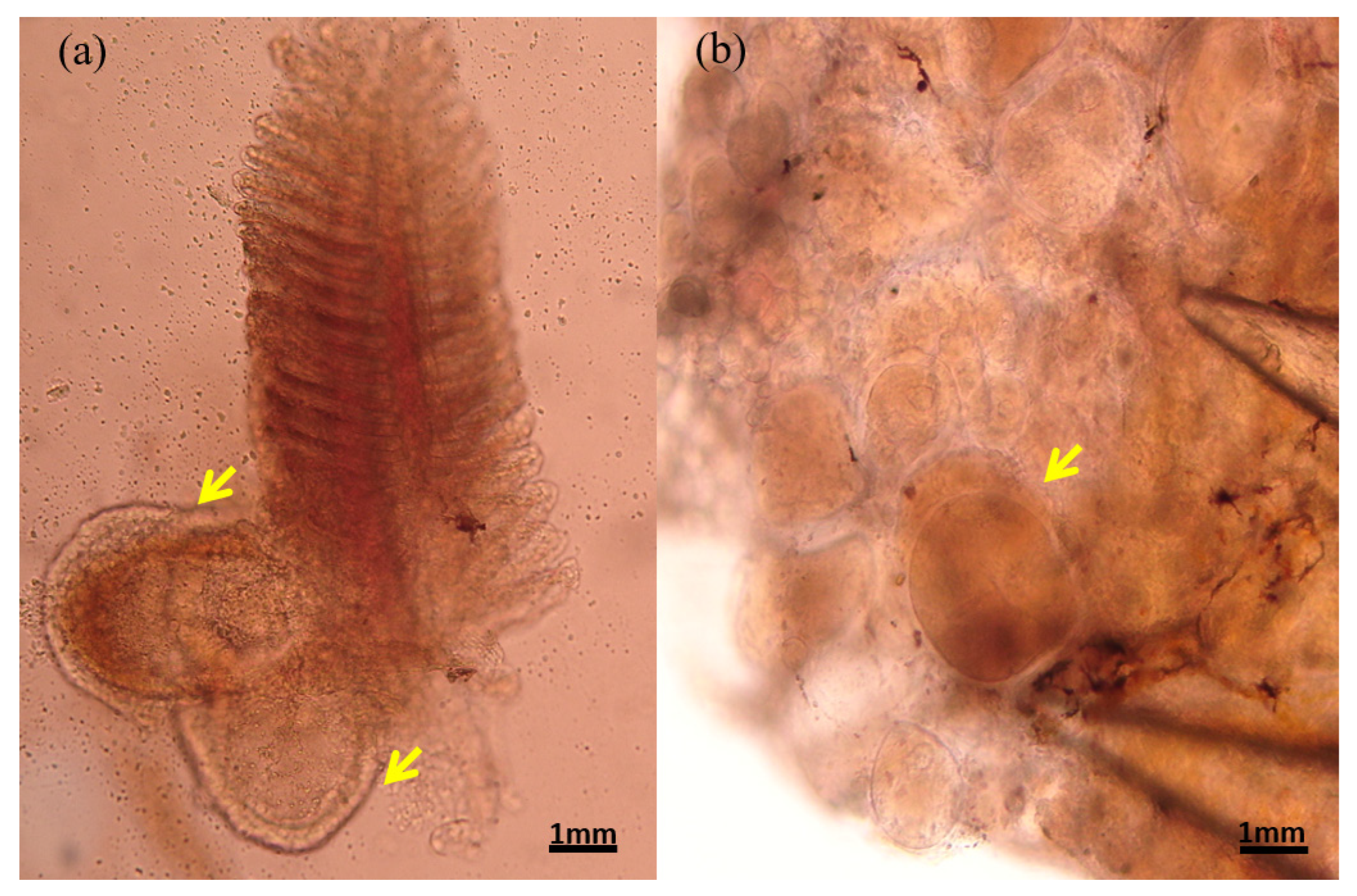
3.1. Macroscopic Observations
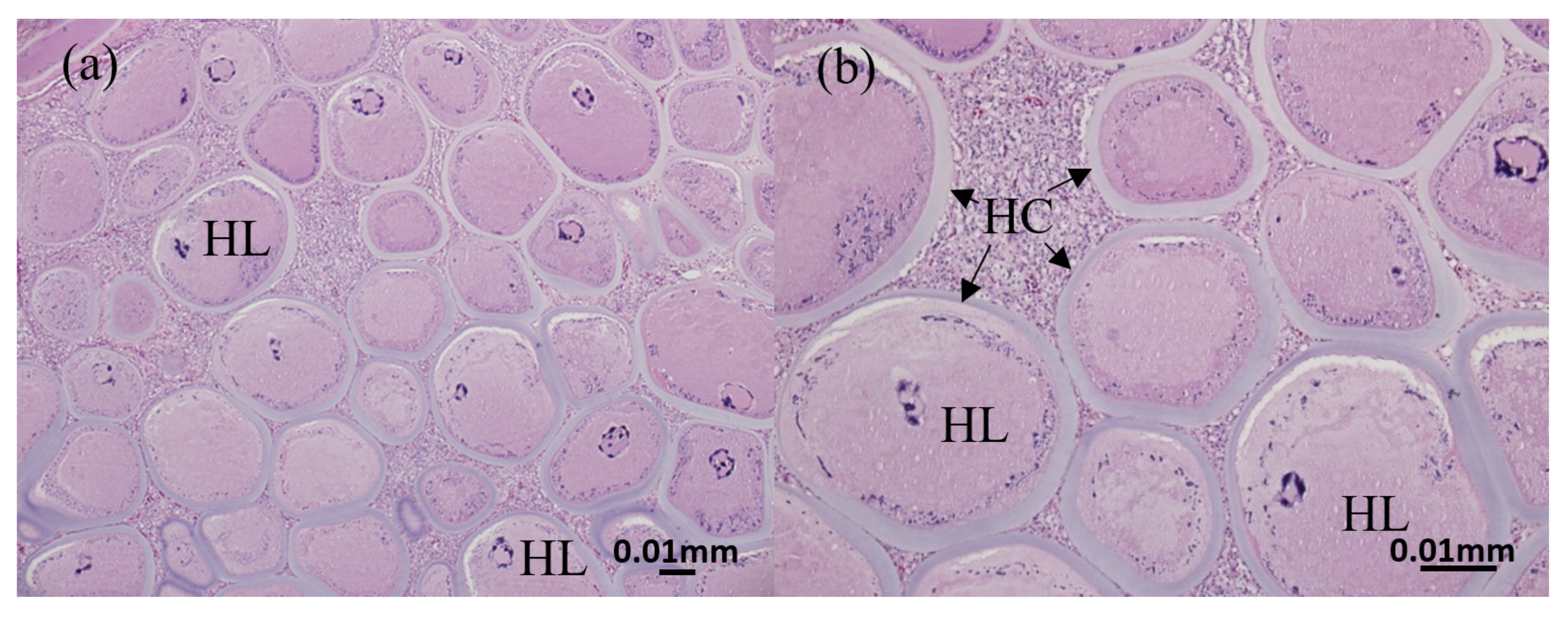
3.2. Histopathology of Nodules from Diseased Clownfish
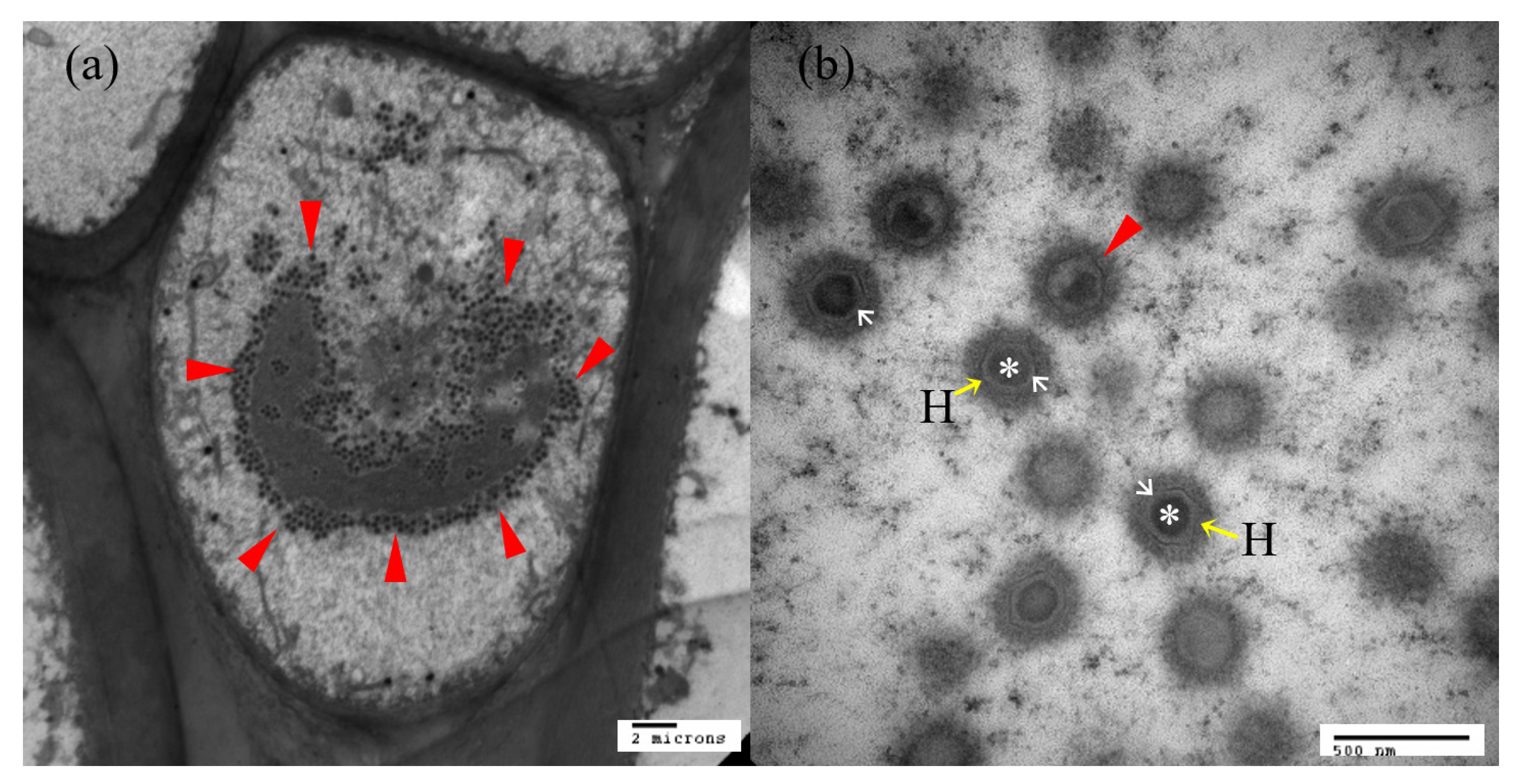
3.3. Electron Microscope
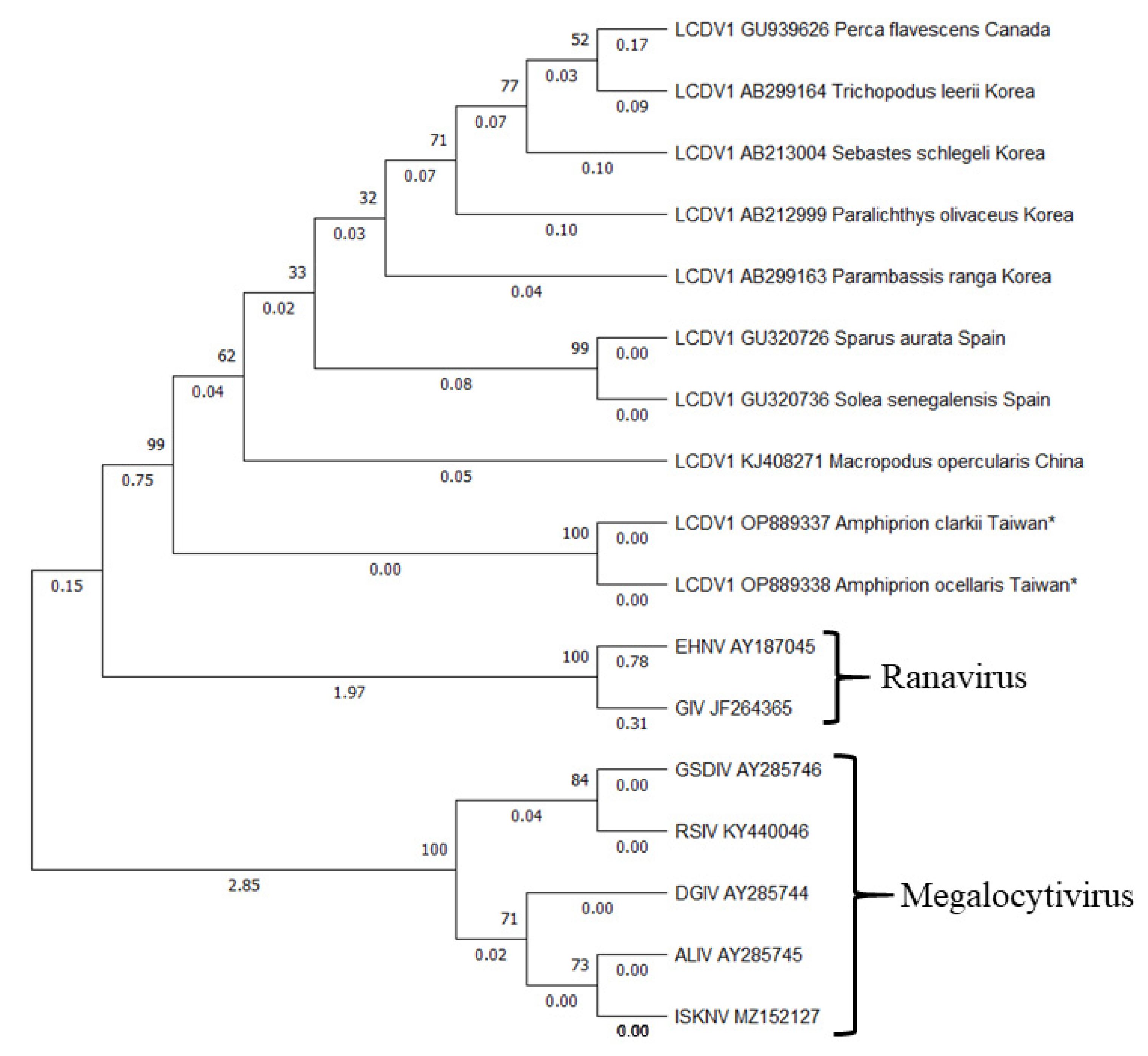
3.4. Molecular Phylogeny
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anders, K. Lymphocystis disease of fishes. In Viruses of Lower Vertebrates; Ahne, W., Kurstak, E., Eds.; Springer: Berlin, Germany, 1989; pp. 140–160. [Google Scholar]
- Wolf, K. Fish Viruses and Fish Viral Disease; Cornell University Press: Ithaca, NY, USA, 1988. [Google Scholar]
- Peter, N.; Schmidt, W. Formation and disintegration of virion in lymphocystis cells of plaice Pleuronectes plassa. Dis. Aquat. Org. 1995, 21, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Chinchar, V.G.; Essbauer, S.M.; He, J.G.; Hyatt, A.; Miyazaki, T.; Seligy, V.; Williams, T. Family Iridoviridae. In Virus Taxonomy: Eighth Report of the International Committee on Taxonomy of Viruses; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberg, U., Ball, L.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 145–162. [Google Scholar]
- Weissenberg, R. Studies on lymphocystis tumor cells of fish; the osmiophilic granules of the cytoplasmic inclusions and their interpretation as elementary bodies of the lymphocystis virus. Cancer Res. 1949, 9, 537–542. [Google Scholar] [PubMed]
- López-Bueno, A.; Mavian, C.; Labella, A.M.; Castro, D.; Borrego, J.J.; Alcami, A.; Alejoc, A. Concurrence of Iridovirus, Polyomavirus, and a unique member of a new group of fish papillomaviruses in lymphocystis disease-affected Gilthead Sea Bream. J Virol. 2016, 90, 8768–8779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirarat, N.; Pratakpiriya, W.; Jongnimitpaiboon, K.; Sajjawiriyakul, K.; Rodkhum, C.; Chansue, N. Lymphocystis disease in cultured false clown anemonefish (Amphiprion ocellaris). Aquaculture 2011, 315, 414–416. [Google Scholar] [CrossRef]
- Xu, L.W.; Feng, J.; Huang, Y.H. Identification of lymphocystis disease virus from paradise fish Macropodus opercularis (LCDV-PF). Arch. Virol. 2014, 159, 2445–2449. [Google Scholar] [CrossRef]
- Tidona, C.A.; Darai, G. The complete DNA sequence of lymphocystis disease virus. Virology 1997, 230, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.Y.; Xiao, F.; Xie, J.; Li, Z.Q.; Gui, J.F. Complete genome sequence of lymphocystis disease virus isolated from China. J Virol. 2004, 78, 6982–6994. [Google Scholar] [CrossRef] [Green Version]
- Doszpoly, A.; Kaján, G.L.; Puentes, R.; Perretta, A. Complete genome sequence and analysis of a novel lymphocystivirusdetected in whitemouth croaker (Micropogonias furnieri): Lymphocystis disease virus 4. Arch. Virol. 2020, 165, 1215–1218. [Google Scholar] [CrossRef] [Green Version]
- Cano, I.; Valverde, E.J.; Lopez-Jimena, B.; Alonso, M.C.; Garcia-Rosado, E.; Sarasquete, C.; Borrego, J.J.; Castro, D. A new genotype of Lymphocystivirus isolated from cultured gilthead seabream, Sparus aurata L., and Senegalese sole, Solea senegalensis (Kaup). J. Fish Dis. 2010, 33, 695–700. [Google Scholar] [CrossRef]
- Palmer, L.J.; Hogan, N.S.; van den Heuvel, M.R. Phylogenetic analysis and molecular methods for the detection of lymphocystis disease virus from yellow perch, Perca flavescens (Mitchell). J. Fish Dis. 2012, 35, 661–670. [Google Scholar] [CrossRef]
- van den Heuvel, M.R.; Power, M.; MacKinnon, M.D.; Van Meer, T.; Dobson, E.P.; Dixon, D.G. Effects of oil sands related aquatic reclamation on yellow perch (Perca flavescens). I. Water quality characteristics and perch physiological and population responses. Can. J. Fish. Aquat. Sci. 1999, 56, 1213–1225. [Google Scholar] [CrossRef]
- Leiva-Rebollo, R.; Labella, A.M.; Valverde, E.J.; Castro, D.; Borrego, J.J. Persistence of Lymphocystis Disease Virus (LCDV) in Seawater. Food Environ. Virol. 2020, 12, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Rahmati-Holasoo, H.; Ahmadivand, S.; Marandi, A.; Shokrpoor, S.; Palić, D.; Jahangard, A. Identification and characterization of lymphocystis disease virus (LCDV) from Indian glassy fish (Parambassis ranga Hamilton, 1822) in Iran. Aquac. Int. 2022, 30, 2593–2602. [Google Scholar] [CrossRef]
- Rahmati-Holasoo, H.; Ghalyanchilangeroudi, A.; Kafi, Z.Z.; Marandi, A.; Shokrpoor, S.; Imantalab, B.; Mousavi, H.E. Detection of lymphocystis disease virus (LCDV) from yellowbar angelfish (Pomacanthus maculosus Forsskål, 1775) in Iran: Histopathological and phylogenetic analysis. Aquaculture 2023, 562, 738862. [Google Scholar] [CrossRef]
- Wabnitz, C.; Taylor, M.; Green, E.; Razak, T. From Ocean to Aquarium; UNEP World Conservation Monitoring Centre: Cambridge, UK, 2003; p. 64. [Google Scholar]
- Abol-Munafi, A.B.; Borazmi-Lokman, N.H.; Asma, N.A.; Sarmiza, S.; Abduh, M.Y. Histological study on the gonad of the protadrous anemonefish (Amphiprion acellaris). J. Anim. Vet. Adv. 2011, 10, 3031–3036. [Google Scholar]
- Cato, J.C.; Brown, C.I. Marine Ornamental Collection, Culture, and Conservation. Open Fish Sci. J. 2003, 2, 55–58. [Google Scholar]
- Siva, U.; Marudhupandi, T.; Haq, M.A.B.; Thankappan, T.; Kumar, A. Histopathological study of lymphocystis disease virus (LCDV) in cultured false clownfish, Amphiprion ocellaris (Cuvier, 1830) and true clownfish, Amphiprion percula (Lacepede, 1802). J. Coast. Life Med. 2014, 2, 264–269. [Google Scholar]
- Lam, C.; Khairunissa, I.; Damayanti, L.; Kurobe, T.; Teh, S.J.; Pfahl, H.; Rapi, S.; Janetski, N.; Baxa, D.V. Detection of a new strain of lymphocystis disease virus (LCDV) in captive-bred clownfish Amphiprion percula in South Sulawesi, Indonesia. Aquac. Int. 2020, 28, 2121–2137. [Google Scholar] [CrossRef]
- Cheng, L.; Lin, W.H.; Wang, P.C.; Tsai, M.A.; Ho, P.Y.; Hsu, J.P.; Chen, R.S.; Chen, S.C. Epidemiology and phylogenetic analysis of Taura syndrome virus in cultured Pacific white shrimp Litopenaeus vannamei B. in Taiwan. Dis. Aquat. Org. 2011, 97, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.; Song, J.; Kitamura, S.; Jung, S.; Oh, M. Phylogenetic analysis of lymphocystis disease virus from tropical ornamental fish species based on a major capsid protein gene. J. Fish Dis. 2008, 31, 473–479. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetic Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Zan, J.D.; Sun, X.Q.; Zhang, Z.W.; Qu, L.Y.; Zhang, J.X. Application of quantitative PCR method in detection of Lymphocystis disease virus China (LCDV-cn) in Japanese flounder (Paralichthys olivaceus). Chin. J. Oceanol. Limnol. 2007, 25, 418–422. [Google Scholar] [CrossRef]
- Cano, I.; Alonso, M.C.; Garcia-Rasado, E.; Saint-Jean, S.R.; Castro, D.; Borrego, J.J. Detection of lymphocystis disease virus (LCDV) in asymptomatic cultured gilt-head seabream (Sparus aurata, L.) using an immunoblot technique. Vet. Microbiol. 2006, 113, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.I.; Jung, S.J.; Kim, W.S.; Nishizawa, T.; Yoshimizu, M.; Oh, M.J. A new genotype of lymphocystivirus, LCDV-RF, from lymphocystis disease rockfish. Arch. Virol. 2006, 151, 607–615. [Google Scholar] [CrossRef]
- Huang, S.M.; Tu, C.; Tseng, C.H.; Huang, C.C.; Kuo, H.C.; Chang, S.K. Genetic analysis of fish iridovirus isolated in Taiwan during 2001–2009. Arch. Virol. 2011, 156, 1505–1515. [Google Scholar] [CrossRef] [Green Version]
- Marsh, I.B.; Whittington, R.J.; O’Rourke, B.; Hyatt, A.D.; Chisholm, O. Rapid differentiation of Australian, European and American ranaviruses based on variation in major capsid protein gene sequence. Mol. Call. Probes. 2002, 16, 137–151. [Google Scholar] [CrossRef]
- Sudthongkong, C.; Miyata, M.; Miyazaki, T. Viral DNA sequences of genes encoding the ATPase and the major capsid protein of tropical iridovirus isolates which are pathogenic to fishes in Japan, South China Sea and Southeast Asian countries. Arch. Virol. 2002, 147, 2089–2109. [Google Scholar] [CrossRef]
- Dong, H.T.; Jitrakorn, S.; Kayansamruaj, P.; Pirarat, N.; Rodkhum, C.; Tattanarojpong, T.; Senapin, S.; Saksmerprome, V. Infectious spleen and kidney necrosis disease (ISKND) outbreaks in farmed barramundi (Lates calcarifer) in Vietnam. Fish Shellfish Immunol. 2017, 86, 65–73. [Google Scholar] [CrossRef]
- Swaminathan, T.R.; Raj, N.S.; Preena, P.G.; Pradhan, P.K.; Sood, N.; Kumar, R.G.; Sudhagar, A.; Sood, N.K. Infectious spleen and kidney necrosis virus-associated large-scale mortality in farmed giant gourami, Osphronemus goramy, in India. J. Fish Dis. 2021, 44, 2043–2053. [Google Scholar] [CrossRef]
- Kawato, S.; Nozaki, R.; Hirono, I.; Kondo, H. Genome Sequence of Lymphocystis Disease Virus 2 LCDV-JP_Oita_2018, Isolated from a Diseased Japanese Flounder (Paralichthys olivaceus) in Japan. Microbiol. Resour. Announc. 2021, 10, e00547-e21. [Google Scholar] [CrossRef]
- Zhan, W.B.; Li, Y.G.; Sheng, X.Z.; Xing, J.; Tang, X.Q. Detection of lymphocystis disease virus in Japanese flounder Paralichthys olivaceus and other marine teleosts from northern China. Chin. J. Oceanol. Limnol. 2010, 28, 1213–1220. [Google Scholar] [CrossRef]
- Sheng, X.Z.; Zheng, W.B.; Xu, S.J.; Cheng, S.F. Histopathological observation of lymphocystis disease and lymphocystis disease virus (LCDV) detection in cultured diseased Sebastes schlegeli. J. Ocean Univ. China 2007, 6, 378–382. [Google Scholar] [CrossRef]
- Zheng, F.R.; Liu, H.Z.; Guo, X.Y.; Wang, B. Isolation and identification of a new isolate of lymphocystis disease virus isolated from black rockfish (Sebastes schlegelii) in China. Aquaculture 2016, 451, 340–344. [Google Scholar] [CrossRef]
- Shawky, M.; Taha, E.; Ahmed, B.; Mahmoud, M.A.; Abdelaziz, M.; Faisal, M.; Yousif, A. Initial Evidence That Gilthead Seabream (Sparus aurata L.) Is a Host for Lymphocystis Disease Virus Genotype, I. Animals 2021, 11, 3032. [Google Scholar] [CrossRef]
- Hassad-Boubaker, S.; Bouzgarou, N.; Fakhfakh, E.; Khayech, M.; Mohamed, S.B.; Megdich, A.; Chéhida, N.B. Detection and genetic characterization of Lymphocystis Disease Virus (LCDV) isolated during disease outbreaks in cultured gilt-head sea bream Sparus aurata in Tunisia. Fish Pathol. 2013, 48, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Alonso, M.C.; Cano, I.; Garcia-Rosado, E.; Cartro, D.; Lamas, J.; Berja, J.L.; Borrego, J.J. Isolation of lymphocystis disease virus from sole, Solea senegalensis Kaup, and blackspot sea bream, Pagellus bogaraveo (Brunnich). J. Fish Dis. 2005, 28, 221–228. [Google Scholar] [CrossRef]
- Kvitt, H.; Heinisch, G.; Diamant, A. Detection and phylogeny of Lymphocystivirus in sea bream Sparus aurata based on the DNA polymerase gene and major capsid protein sequences. Aquaculture 2008, 275, 58–63. [Google Scholar] [CrossRef]
- Carballo, C.; Castro, D.; Borrego, J.J.; Manchado, M. Gene expression profiles associated with lymphocystis disease virus (LCDV) in experimentally infected Senegalese sole (Solea senegalensis). Fish Shellfish Immunol. 2017, 66, 129–139. [Google Scholar] [CrossRef]
- Hu, G.B.; Lou, H.M.; Dong, X.Z.; Liu, Q.M.; Zhang, S.C. Characteristics of the interferon regulatory factor 5 (IRF5) and its expression in response to LCDV and poly I: C challenges in Japanese flounder, Paralichthys olivaceus. Dev. Comp. Immunol. 2012, 38, 377–382. [Google Scholar] [CrossRef]
- Valverde, E.J.; Borrego, J.J.; Sarasquete, M.C.; Ortiz-Delgado, J.B.; Castro, D. Target organs for lymphocystis disease virus replication in gilthead seabream (Sparus aurata). Vet. Res. 2017, 48, 21. [Google Scholar] [CrossRef] [Green Version]
- Sheng, X.Z.; Zeng, J.; Zhong, Y.; Tang, X.Q.; Xing, J.; Chi, H.; Zhan, W.B. Peripheral blood B-lymphocytes are involved in lymphocystis disease virus infection in flounder (Paralichthys olivaceus) via cellular receptor-mediated mechanism. Int. J. Mol. Sci. 2022, 23, 9225. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Code | Virus Species | Host | Region | Size | References |
|---|---|---|---|---|---|
| OP889337 | LCDV 1 | Amphiprion clarkii | Taiwan | 1390 | Present study |
| OP889338 | LCDV 1 | Amphiprion ocellaris | Taiwan | 1390 | Present study |
| AB299163 | LCDV 1 | Parambassis ranga | Korea | 1337 | [26] |
| KJ408271 | LCDV 1 | Macropodus opercularis | China | 1208 | [8] |
| GU320726 | LCDV 1 | Sparus aurata | Spain | 1317 | [12] |
| GU320736 | LCDV 1 | Solea senegalensis | Spain | 1317 | [27] |
| AB212999 | LCDV 1 | Paralichthys olivaceus | Korea | 1347 | [28] |
| GU939626 | LCDV 1 | Perca flavescens | Canada | 1357 | [13] |
| AB299164 | LCDV 1 | Trichopodus leerii | Korea | 1337 | [24] |
| AB213004 | LCDV 1 | Sebastes schlegeli | Korea | 1356 | [28] |
| JF264365 | GIV | Epinephelus coioides | Taiwan | 1392 | [29] |
| AY187045 | EHNV | Perca fluviatilis | Australia | 1472 | [30] |
| AY285746 | GSDIV | Epinephelus chlorostigma | Thailand | 1362 | [31] |
| KY440046 | RSIV | Lates calcarifer | Vietnam | 1362 | [32] |
| AY285745 | ALIV | Aplocheilichthys centralis | Indonesia | 1362 | [31] |
| MZ152127 | ISKNV | Osphronemus goramy | India | 1362 | [33] |
| AY285744 | DGIV | Colisa lalia | Malaysia | 1362 | [31] |
| Clownfish Species | Organ | ||
|---|---|---|---|
| Gill | Fin | Spleen | |
| Amphiprion clarkii (n = 1) | + | + | - |
| Amphiprion ocellaris (n = 4) | + | + | + |
| No. | Accession Number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | OP889337/ OP889338 | 100.0 | 92.5 | 91.5 | 90.8 | 90.5 | 83.4 | 83.0 | 81.3 | 80.5 | 57.3 | 55.9 | 55.9 | 55.9 | 55.9 | 55.7 | 55.7 |
| 2 | KJ408271 | 100.0 | 89.9 | 89.6 | 89.2 | 86.0 | 83.8 | 83.9 | 83.3 | 56.6 | 52.8 | 54.4 | 54.2 | 54.2 | 54.1 | 54.2 | |
| 3 | AB299163 | 100.0 | 89.1 | 88.8 | 86.1 | 82.9 | 84.3 | 85.2 | 55.7 | 52.2 | 54.2 | 53.7 | 53.5 | 53.5 | 54.1 | ||
| 4 | GU320726 | 100.0 | 99.7 | 84.8 | 83.4 | 83.2 | 84.0 | 56.1 | 53.6 | 54.8 | 54.1 | 54.1 | 54.1 | 54.7 | |||
| 5 | GU320736 | 100.0 | 84.5 | 83.1 | 82.9 | 83.7 | 55.9 | 53.6 | 54.7 | 54.0 | 54.0 | 54.0 | 54.5 | ||||
| 6 | AB212999 | 100.0 | 82.3 | 84.8 | 85.6 | 56.3 | 50.8 | 52.2 | 51.9 | 51.7 | 51.6 | 52.2 | |||||
| 7 | GU939626 | 100 | 84.6 | 83.5 | 55.8 | 51.0 | 53.8 | 53.3 | 53.3 | 53.1 | 53.7 | ||||||
| 8 | AB299164 | 100.0 | 87.4 | 54.9 | 49.2 | 52.7 | 52.3 | 52.2 | 52.1 | 52.7 | |||||||
| 9 | AB213004 | 100.0 | 54.3 | 49.9 | 52.4 | 51.1 | 50.9 | 50.9 | 52.2 | ||||||||
| 10 | GIV | 100.0 | 69.9 | 50.7 | 50.7 | 50.9 | 50.5 | 50.5 | |||||||||
| 11 | EHNV | 100.0 | 54.6 | 54.5 | 54.5 | 54.3 | 54.7 | ||||||||||
| 12 | RSIV | 100.0 | 95.0 | 94.6 | 94.9 | 99.4 | |||||||||||
| 13 | ALIV | 100.0 | 99.7 | 99.7 | 94.7 | ||||||||||||
| 14 | ISKNV | 100.0 | 99.3 | 94.4 | |||||||||||||
| 15 | DGIV | 100.0 | 94.6 | ||||||||||||||
| 16 | GSDIV | 100.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, M.-C.; See, M.S.; Wang, P.-C.; Kuo, Y.-T.; Ho, Y.-S.; Chen, S.-C.; Tsai, M.-A. Lymphocystis Disease Virus Infection in Clownfish Amphiprion ocellaris and Amphiprion clarkii in Taiwan. Animals 2023, 13, 153. https://doi.org/10.3390/ani13010153
Cheng M-C, See MS, Wang P-C, Kuo Y-T, Ho Y-S, Chen S-C, Tsai M-A. Lymphocystis Disease Virus Infection in Clownfish Amphiprion ocellaris and Amphiprion clarkii in Taiwan. Animals. 2023; 13(1):153. https://doi.org/10.3390/ani13010153
Chicago/Turabian StyleCheng, Ming-Chung, Ming She See, Pei-Chi Wang, Yu-Ting Kuo, Yuan-Shing Ho, Shih-Chu Chen, and Ming-An Tsai. 2023. "Lymphocystis Disease Virus Infection in Clownfish Amphiprion ocellaris and Amphiprion clarkii in Taiwan" Animals 13, no. 1: 153. https://doi.org/10.3390/ani13010153