
Morphological and Molecular Identification of Mullet Helminth Parasite Fauna from Ganzirri Lagoon (Sicily, Southern Italy)
 , ,
, ,  ,
,  ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Sampling
2.2. Parasitological Examination
2.3. Molecular Analysis
2.4. Statistical Analysis
3. Results
3.1. Parasitological Findings
3.2. Molecular Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holmlund, C.M.; Hammer, M. Ecosystem services generated by fish populations. Ecol. Econom. 1999, 29, 253–268. [Google Scholar] [CrossRef]
- Yilmaz, F. The comparison of heavy metal concentrations (Cd, Cu, Mn, Pb, and Zn) in tissues of three economically important fish (Anguilla anguilla, Mugil cephalus and Oreochromis niloticus) inhabiting Koycegiz Lake-Mugia (Turkey). Turk. J. Sci. Technol. 2009, 4, 7–15. [Google Scholar]
- Wilson, R.W.; Millero, F.J.; Taylor, J.R.; Walsh, P.J.; Christensen, V.; Jennings, S.; Grosell, M. Contribution of fish to the marine inorganic carbon cycle. Science 2009, 323, 359–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balk, L.; Larsson, A.; Frolin, L. Baseline studies of biomarkers in the feral female perch (Perca fluviatilis) as tools in biological monitoring of anthropogenic substances. Mar. Environ. Res. 1996, 42, 203–208. [Google Scholar] [CrossRef]
- Curson, A.R.J.; Matthew, J.S.; Todd, J.D.; Carpenter, S.R.; Cottingham, K.L. Resilience and restoration of lakes. Conserv. Ecol. 1997, 1, 2. [Google Scholar]
- Diop, M.; Howsam, M.; Diop, C.; Goossens, J.F.; Diouf, A.; Amara, R. Assessment of trace element contamination and bioaccumulation in algae (Ulva lactuca), mussels (Perna perna), shrimp (Penaeus kerathurus), and fish (Mugil cephalus, Saratherondon melanotheron) along the Senegalese coast. Mar. Pollut. Bull. 2016, 103, 339–343. [Google Scholar] [CrossRef]
- Ihunwo, O.C.; Dibofori-Orji, A.N.; Olowu, C.; Ibezim-Ezeani, M.U. Distribution and risk assessment of some heavy metals in surface water, sediment and grey mullet (Mugil cephalus) from contaminated creek in Woji, southern Nigeria. Mar. Poll. Bull. 2020, 154, 111042. [Google Scholar] [CrossRef]
- Crosetti, D. Biology, Ecology and Culture of Grey mullet (Mugilidae), 1st ed.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2016; pp. 42–127. [Google Scholar]
- Míguez-Lozano, R.; Pardo-Carranza, T.V.; Blasco-Costa, I.; Balbuena, J.A. Spatial structure of helminth communities in the golden grey mullet, Liza aurata (Actinopterygii: Mugilidae), from the Western Mediterranean. J. Parasitol. 2012, 98, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Llopis-Belenguer, C.; Pavoine, S.; Blasco-Costa, I.; Balbuena, J.A. Assembly rules of helminth parasite communities in grey mullets: Combining components of diversity. Int. J. Parasitol. 2020, 50, 1089–1098. [Google Scholar] [CrossRef]
- Christison, K.W.; Vaughan, D.B.; Shinn, A.P.; Hansen, H. Gyrodactylus molweni sp. n. (Monogenea: Gyrodactylidae) from Chelon richardsonii (Smith, 1846) (Mugilidae) from Table Bay, South Africa. Int. J. Parasitol. Parasites Wildl 2021, 15, 87–94. [Google Scholar] [CrossRef]
- Ovcharenko, M. Microparasites of worldwide mullets. Ann. Parasitol. 2015, 61, 229–239. [Google Scholar] [CrossRef]
- Morales Martínez, K.A.; Muñoz García, C.; Figueroa Delgado, A.; Chávez Güitrón, L.; Osorio Saravia, D.; Saavedra Montañez, M.; Martínez Maya, J.J.; Rubio, J.M.; Villalobos, N. Parasite identification in mullet fish (Mugil curema) from Chautengo Lagoon, Guerrero, Mexico, based on morphology and molecular analysis. Exp. Parasitol. 2022, 240, 108335. [Google Scholar] [CrossRef] [PubMed]
- González-Castro, M.; Macchi, G.J.; Cousseau, M.B. Studies on reproduction of the mullet Mugil platanus Günther, 1880 (Actinopterygii, Mugilidae) from the Mar Chiquita coastal lagoon, Argentina: Similarities and differences with related species. Ital. J. Zool. 2011, 78, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Hastings, P.A. Complementary approaches to systematic ichthyology. Zootaxa 2011, 2946, 57–59. [Google Scholar] [CrossRef]
- Bottari, A.; Bottari, C.; Carveni, P.; Giacobbe, S.; Spanò, N. Genesis and Geomorphologic and Ecological Evolution of the Ganzirri Salt Marsh (Messina, Italy). Quat. Int. 2005, 140–141, 150–158. [Google Scholar] [CrossRef]
- Paperna, I.; Lahav, M. New records and further data on fish parasites in Israel. Bamidgeh 1971, 23, 43–52. [Google Scholar]
- Paperna, I. Parasites and diseases of the grey mullet (Mugilidae) with special reference to the seas of the Near East. Aquaculture 1975, 5, 65–80. [Google Scholar] [CrossRef]
- Matthews, R.A.; Burgess, P.J. Cryptocaryon Irritans (Ciliophora): Primary Infection in Thick-Lipped Mullet, Chelon Labrosus (Risso). J. Fish Dis. 1995, 18, 329–335. [Google Scholar] [CrossRef]
- Wilkie, D.W. Outbreak of cryptocaryoniasis in marine aquaria at Scripps Institute of Oceanography. Calif. Fish Game 1969, 55, 227–236. [Google Scholar]
- Özer, A.; Öztürk, T. Trichodina puytoraci Lom, 1962 and Trichodina lepsii Lom, 1962 (Peritrichida: Ciliophora) infestations on Mugilids caught at the Black Sea coast of Sinop in Turkey. Turk. J. Zool. 2004, 28, 179–182. [Google Scholar]
- Marcotegui, P.S.; Martorelli, S.R. Trichodinids (Ciliophora: Peritrichida) of Mugil platanus (Mugiliformes: Mugilidae) and Micropogonias furnieri (Perciformes: Sciaenidae) from Samborombón Bay, Argentina, with the description of a new species. Folia Parasitol. 2009, 56, 167–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigrelli, R.F. Enzootics in the New York Aquarium caused by Cryptocaryon irritans Brown, 1951 (=Ichthyophthirius marinus Sikama, 1961), a histophagous ciliate in the skin, eyes and gills of marine fishes. Zoologica 1966, 51, 97–102. [Google Scholar] [CrossRef]
- Domnich, I.F.; Sarabeev, V.L. Formation of the parasite fauna of Mugil soiuy in the Azov Sea. Visnyk Zaporizkogo Derzhavnogo Universytetu 1999, 2, 218–223. [Google Scholar]
- Lawler, A.R. Studies on Amyloodinium ocellatum (Dinoflagellata) in Mississippi Sound: Natural and experimental hosts. Gulf and Carib. Res. 1980, 6, 403–413. [Google Scholar] [CrossRef]
- Baticados, M.C.L.; Quinitio, G.F. Occurrence and pathology of an Amyloodinium-like protozoan parasite on gills of grey mullet, Mugil cephalus. Helgoländer Meeresunters. 1984, 37, 595–601. [Google Scholar]
- Merella, P.; Garippa, G. Metazoan parasites of Grey mullets (Teleostea: Mugilidae) from the Mistras Lagoon (Sardinia-western Mediterranean). Sci. Mar. 2001, 65, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Masala, S.; Piras, M.C.; Sanna, D.; Chai, J.Y.; Jung, B.K.; Sohn, W.M.; Garippa, G.; Merella, P. Epidemiological and molecular data on heterophyid trematode metacercariae found in the muscle of Grey mullets (Osteichthyes: Mugilidae) from Sardinia (western Mediterranean Sea). Parasitol. Res. 2016, 115, 3409–3417. [Google Scholar] [CrossRef] [PubMed]
- Scholz, T.; Aguirre-Macedo, M.L.; Salgado-Maldonado, G. Trematodes of the family Heterophyidae (Digenea) in Mexico: A review of species and new host and geographical records. J. Nat. Hist. 2001, 35, 1733–1772. [Google Scholar] [CrossRef]
- Dmitrieva, E.V.; Gaevskaya, A.V. Parasitological aspects of mugilids mariculture and of their introduction into the Sea of Azov and the Black Sea. Ekol. Morya Kiev 2001, 55, 73–78. [Google Scholar]
- Oguz, M.C.; Bray, R.A. Cestoda and Monogenea of some teleost fishes off the Mudanya Coast (Sea of Marmara, Turkey). Helminthologia 2008, 45, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Caltran, H.; Silan, P. Gill Filaments of Liza Ramada, a Biotope for Ectoparasites: Surface Area Acquisition Using Image Analysis and Growth Models. J. Fish Biol. 1996, 49, 1267–1279. [Google Scholar] [CrossRef]
- Blasco-Costa, I.; Balbuena, J.A.; Raga, J.A.; Kostadinova, A.; Olson, P.D. Molecules and morphology reveal cryptic variation among digeneans infecting sympatric mullets in the Mediterranean. Parasitology 2010, 137, 287–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jithendran, K.P.; Kannappan, S. A short note on heavy infection of acanthocephalan worm (Neoechinorhynchus agilis) in Grey mullet, Mugil cephalus. J. Parasit. Dis. 2010, 34, 99–101. [Google Scholar] [CrossRef] [Green Version]
- Ragias, V.; Athanassopoulou, F.; Sinis, A. Parasites of Mugilidae spp. reared under semi-intensive and intensive conditions in Greece. Bull. Eur. Assoc. Fish Pathol. 2005, 25, 107–113. [Google Scholar]
- Castellanos, J.A.; Tangua, A.R.; Salazar, L. Anisakidae nematodes isolated from the flathead grey mullet fish (Mugil cephalus) of Buenaventura, Colombia. Int. J. Parasitol. Parasites Wildl. 2017, 6, 265–270. [Google Scholar] [CrossRef]
- Valles-Vega, I.; Molina-Fernández, D.; Benítez, R.; Hernández-Trujillo, S.; Adroher, F.J. Early development and life cycle of Contracaecum multipapillatum s.l. from a brown pelican Pelecanus occidentalis in the Gulf of California, Mexico. Dis. Aquat. Org. 2017, 125, 167–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamsi, S.; Suthar, J. A revised method of examining fish for infection with zoonotic nematode larvae. Int. J. Food Microbiol. 2016, 227, 13–16. [Google Scholar] [CrossRef]
- Petter, A.J.; Maillard, C. Larves d’ascarides parasites de poissons en Méditerranée occidentale. Bull. Mus. Natl. Hist. Nat. 1988, 10, 347–369. [Google Scholar]
- Yamaguti, S. Digenetic Trematodes of Hawaiian Fishes; Interscience Publishers: New York, NY, USA, 1970. [Google Scholar]
- Tkach, I.V.; Sarabeev, V.L.; Shvetsova, L.S. Taxonomic status of Neoechinorhynchus agilis (Acanthocephala, Neoechinorhynchidae), with a description of two new species of the genus from the Atlantic and Pacific mullets (Teleostei, Mugilidae). Vestnik Zoologii 2014, 48, 291–306. [Google Scholar] [CrossRef] [Green Version]
- Routtu, J.; Grunberg, D.; Izhar, R.; Dagan, Y.; Guttel, Y.; Ucko, M.; Ben-Ami, F. Selective and universal primers for trematode barcoding in freshwater snails. Parasitol. Res. 2014, 113, 2535–2540. [Google Scholar] [CrossRef]
- Mladineo, I.; Bott, N.J.; Nowak, B.F.; Block, B.A. Multilocus phylogenetic analyses reveal that habitat selection drives the speciation of Didymozoidae (Digenea) parasitizing Pacific and Atlantic bluefin tunas. Parasitology 2010, 137, 1013–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology Meets Ecology on Its Own Terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Sayyaf Dezfuli, B.; Castaldelli, G.; Giari, L. Histopathological and ultrastructural assessment of two mugilid species infected with myxozoans and helminths. J. Fish Dis. 2018, 41, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Paperna, I.; Dzikowski, R. Digenea (phylum Platyhelminthes). In Fish Diseases and Disorders; Volume 1: Protozoan and metazoan infections; Woo, P.T.K., Ed.; CAB International: Wallingford, CT, USA, 2006; pp. 345–390. [Google Scholar]
- Amin, O.M.; Sharifdini, M.; Heckmann, R.A.; Rubtsova, N.; Chine, H.J. On the Neoechinorhynchus agilis (Acanthocephala: Neoechinorhynchidae) complex, with a description of Neoechinorhynchus ponticus n. sp. from Chelon auratus in the Black Sea. Parasite 2020, 27, 48. [Google Scholar] [CrossRef]
- Palombi, A. Per una migliore conoscenza del Trematodi endoparassiti dei pesci del golfo di Napoli. I. Steringotrema divergens (Rud.) e Haploporus benedeni (Stoss.). Annuario del Museo Zoologico R Uni Napoli 1931, 6, 1–15. [Google Scholar]
- Ergens, R. Helminth fauna of some fish in Albania. Ceskoslovenská parasitologie 1960, 7, 49–90. [Google Scholar]
- Fares, A.; Maillard, C. Recherches sur quelques Haploporidae (Trematoda) parasites des Muges de Méditerranée Occidentale: Systématique et cycles évolutifs. Zeit Für Pare 1974, 45, 11–43. [Google Scholar]
- Radujković, B.M.; Raibaut, A. Parasites des poissons marins du Montenegro: Liste des espèces de poissons avec leurs parasites. Acta Adriatica 1989, 30, 307–319. [Google Scholar]
- Saad-Fares, A.; Maillard, C. Étude en microscopie électronique à balayage du kyste métacercarien de Saccocoelium tensum Looss, 1902 (Trematoda-Haploporidae). Ann. Parasitol. Hum. Comp. 1985, 60, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Al-Bassel, D.A.; Hussein, A.N.A. A survey on parasites infecting mullets from Egypt and Libya. Egypt. Acad. J. Biol. Sci. 2012, 4, 9–19. [Google Scholar] [CrossRef]



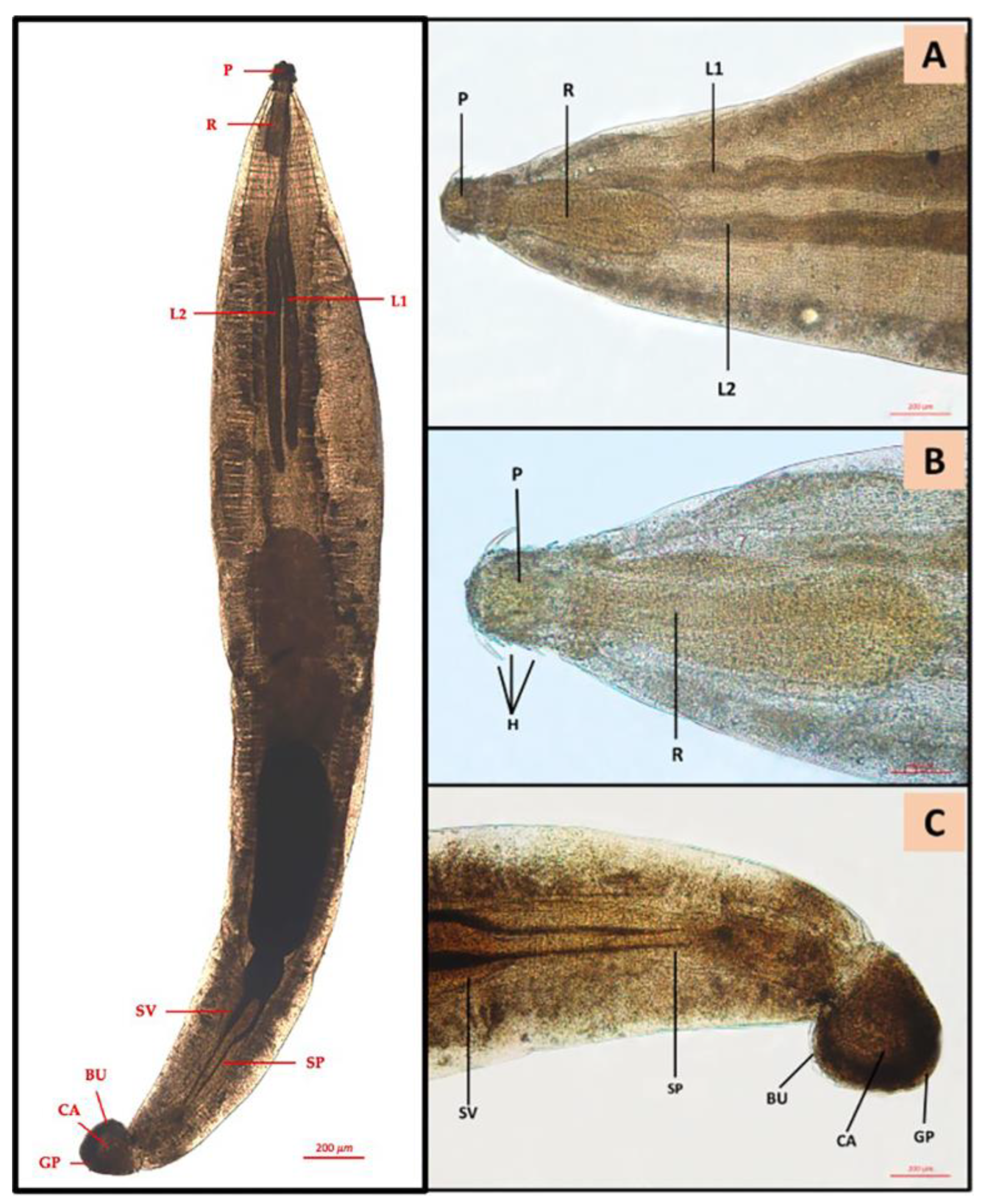{kind=link}
{kind=link}
| Species | Mean Length (cm) ± SD | Mean Weight (g) ± SD |
|---|---|---|
| Chelon labrosus | 20.18 ± 3.99 | 91.66 ± 139.67 |
| Chelon auratus | 19.75 ± 1.9 | 72.08 ± 22.19 |
| Oedalechilus labeo | 18.75 ± 2.74 | 65.28 ± 23.97 |
| Chelon labrosus | |||
| S | I | GIT | |
| Prevalence % | 12.12 | 14.14 | 24.24 |
| Mean intensity | 6.16 | 7.78 | 346.3 |
| Mean abundance | 0.74 | 1.1 | 83.9 |
| Chelon auratus | |||
| S | I | GIT | |
| Prevalence % | 2.7 | 16.2 | 8.1 |
| Mean intensity | 2 | 7 | 57.6 |
| Mean abundance | 0.05 | 1.13 | 4.67 |
| Oedalechilus labeo | |||
| S | I | GIT | |
| Prevalence % | 50 | - | 50 |
| Mean intensity | 8.66 | - | 6.5 |
| Mean abundance | 1.85 | - | 1.85 |
| Kruskal–Wallis Test | |||
| p value | 0.0374 | ||
| p value summary | * | ||
| Do the medians vary signif. (p < 0.05)? | Yes | ||
| Number of groups | 3 | ||
| Kruskal–Wallis statistic | 6.573 | ||
| Dunn’s multiple comparisons test | Mean rank diff. | Summary | Adjusted p Value |
| H. benedeni -CA vs. H. benedeni -CL | −19.29 | * | 0.0343 |
| H. benedeni -CA vs. H. benedeni -OL | −18.69 | ns | 0.397 |
| H. benedeni -CL vs. H. benedeni -OL | 0.5988 | ns | >0.9999 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Benedetto, G.; Capparucci, F.; Natale, S.; Savoca, S.; Riolo, K.; Gervasi, C.; Albano, M.; Giannetto, A.; Gaglio, G.; Iaria, C. Morphological and Molecular Identification of Mullet Helminth Parasite Fauna from Ganzirri Lagoon (Sicily, Southern Italy). Animals 2023, 13, 847. https://doi.org/10.3390/ani13050847
De Benedetto G, Capparucci F, Natale S, Savoca S, Riolo K, Gervasi C, Albano M, Giannetto A, Gaglio G, Iaria C. Morphological and Molecular Identification of Mullet Helminth Parasite Fauna from Ganzirri Lagoon (Sicily, Southern Italy). Animals. 2023; 13(5):847. https://doi.org/10.3390/ani13050847
Chicago/Turabian StyleDe Benedetto, Giovanni, Fabiano Capparucci, Sabrina Natale, Serena Savoca, Kristian Riolo, Claudio Gervasi, Marco Albano, Alessia Giannetto, Gabriella Gaglio, and Carmelo Iaria. 2023. "Morphological and Molecular Identification of Mullet Helminth Parasite Fauna from Ganzirri Lagoon (Sicily, Southern Italy)" Animals 13, no. 5: 847. https://doi.org/10.3390/ani13050847