
On the Species Identification of Two Non-Native Tilapia Species, Including the First Record of a Feral Population of Oreochromis aureus (Steindachner, 1864) in South Korea

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Classification according to the Morphological Identification
2.3. Identification
2.4. Mitochondrial DNA Sequencing
2.5. Genetic Data Analysis
3. Results
3.1. Photographic Record
3.2. Morphological Species Identification
3.3. Molecular Identification
3.4. Presumption of the Tilapia Introduction Area
4. Discussion
4.1. Comparison of Morphological Analyses Results
4.2. Comparison of Molecular Analyses Results
4.3. Potential Impact of the Aquatic Ecosystem
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McAndrew, B.J. Evolution, phylogenetic relationships and biogeography. In Tilapias: Biology and Exploitation; McAndrew, B.J., Ed.; Springer: Berlin, Germany, 2000; Volume 25, pp. 1–32. [Google Scholar]
- Trewavas, E. Tilapiine Fishes of the Genera Sarotherodon, Oreochromis and Danakilia; British Museum (Natural History): London, UK, 1983; p. 583. [Google Scholar]
- FAO. Towards Blue Transformation. In The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2022; p. 266. [Google Scholar]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- FAO. Opportunities and challenges. In The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2014; p. 1243. [Google Scholar]
- Kim, D.S.; Park, I.-S. Genetic identification of hatchery reared tilapia strains. J. Aquac. 1990, 3, 31–37. [Google Scholar]
- Courtenay, W. Aquaculture in the Americas. In Tilapias as Non-Indigenous Species in the Americas: Environmental, Regulatory and Legal Issues; World Aquaculture Society: Sorrento, LA, USA, 1997; Volume 1, pp. 18–33. [Google Scholar]
- Costa-Pierce, B.A. Rapid evolution of an established feral tilapia (Oreochromis spp.): The need to incorporate invasion science into regulatory structures. Biol. Invasions. 2003, 5, 71–84. [Google Scholar] [CrossRef]
- Clavero, M.; García-Berthou, E. Homogenization dynamics and introduction routes of invasive freshwater fish in the Iberian Peninsula. Ecol. Appl. 2006, 16, 2313–2324. [Google Scholar] [CrossRef] [Green Version]
- Gozlan, R.E.; Britton, J.; Cowx, I.; Copp, G. Current knowledge on non-native freshwater fish introductions. J. Fish Biol. 2010, 76, 751–786. [Google Scholar] [CrossRef]
- Canonico, G.C.; Arthington, A.; McCrary, J.K.; Thieme, M.L. The effects of introduced tilapias on native biodiversity. Aquat. Conserv. Mar. Freshw. 2005, 15, 463–483. [Google Scholar] [CrossRef]
- Casal, C.M.V. Global documentation of fish introductions: The growing crisis and recommendations for action. Biol. Invasions. 2006, 8, 3–11. [Google Scholar] [CrossRef]
- Vicente, I.; Fonseca-Alves, C. Impact of introduced Nile tilapia (Oreochromis niloticus) on non-native aquatic ecosystems. Pak. J. Biol. Sci. 2013, 16, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Lèveque, C. Out of Africa: The Success Story of Tilapias. Environ. Biol. Fishes. 2002, 64, 461–464. [Google Scholar] [CrossRef]
- de Attayde, J.L.; Okun, N.; Brasil, J.; Menezes, R.; Mesquita, P. Impactos da introdução da tilápia do Nilo, Oreochromis niloticus, sobre a estrutura trófica dos ecossistemas aquáticos do Bioma Caatinga. Oecol. Aust. 2007, 11, 450–461. [Google Scholar] [CrossRef]
- Charo-Karisa, H.; Rezk, M.A.; Bovenhuis, H.; Komen, H. Heritability of cold tolerance in Nile tilapia, Oreochromis niloticus, juveniles. Aquac. Res. 2005, 249, 115–123. [Google Scholar] [CrossRef]
- Zhu, H.; Liu, Z.; Lu, M.; Gao, F.; Ke, X.; Ma, D.; Huang, Z.; Cao, J.; Wang, M. Screening and identification of a microsatellite marker associated with sex in Wami tilapia, Oreochromis urolepis hornorum. J. Genet. 2016, 95, 283–289. [Google Scholar] [CrossRef]
- Kong, D.; Lee, O.; Song, H.; Song, M. Survey and Recovery Measures on the Aquatic Ecosystem of Jinwi Stream; Gyeonggi Development Institute: Goyang, Korea, 2011; pp. 1–145. [Google Scholar]
- Yoon, J.-D.; Kim, J.-H.; Park, S.-H.; Jang, M.-H. The distribution and diversity of freshwater fishes in Korean Peninsula. J. Ecol. Environ. 2018, 51, 71–85. [Google Scholar] [CrossRef]
- Wang, J.H.; Choi, J.K.; Lee, H.G. The population characteristics of Nile tilapia (Oreochromis niloticus) in Dalseo Stream, South Korea. Korean J. Environ. Biol. 2020, 38, 127–136. [Google Scholar] [CrossRef]
- Padilla, D.K.; Williams, S.L. Beyond ballast water: Aquarium and ornamental trades as sources of invasive species in aquatic ecosystems. Front. Ecol. Environ. 2004, 2, 131–138. [Google Scholar] [CrossRef]
- Anane-Taabeah, G.; Frimpong, E.A.; Hallerman, E. Aquaculture-mediated invasion of the genetically improved farmed tilapia (Gift) into the Lower Volta Basin of Ghana. Diversity 2019, 11, 188. [Google Scholar] [CrossRef] [Green Version]
- Maranan, J.B.D.; Basiao, Z.U.; Quilang, J.P. DNA barcoding of feral tilapias in Philippine lakes. Mitochondrial DNA A 2016, 27, 4302–4313. [Google Scholar] [CrossRef]
- Wohlfarth, G.; Hulata, G. Applied Genetics of Tilapias. In Manila (Philippines): ICLARM Studies and Reviews 6, 2nd ed.; International Center for Living Aquatic Resources Management: Metro Man, Philippines, 1981; Volume 6, pp. 1–26. [Google Scholar]
- D’Amato, M.E.; Esterhuyse, M.M.; Van Der Waal, B.C.; Brink, D.; Volckaert, F.A. Hybridization and phylogeography of the Mozambique tilapia Oreochromis mossambicus in southern Africa evidenced by mitochondrial and microsatellite DNA genotyping. Conserv. Genet. 2007, 8, 475–488. [Google Scholar] [CrossRef]
- Syaifudin, M.; Bekaert, M.; Taggart, J.B.; Bartie, K.L.; Wehner, S.; Palaiokostas, C.; Khan, M.; Selly, S.-L.C.; Hulata, G.; D’Cotta, H. Species-specific marker discovery in Tilapia. Sci. Rep. 2019, 9, 13001. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Cao, Z.-D.; Fu, S.-J. The effects of caudal fin loss and regeneration on the swimming performance of three cyprinid fish species with different swimming capacities. J. Exp. Biol. 2013, 216, 3164–3174. [Google Scholar] [CrossRef] [Green Version]
- Valikhani, H.; Abdoli, A.; Kiabi, B.H.; Nejat, F. First record and distribution of the blue tilapia, Oreochromis aureus (Steindachner, 1864) (Perciformes: Cichlidae) in inland waters of Iran. Iran. J. Ichthyol. 2016, 3, 19–24. [Google Scholar] [CrossRef]
- Nico, L.G.; Fuller, P.; Neilson, M.E. Oreochromis aureus (Steindachner, 1864); U.S. Geological Survey, Nonindigenous Aquatic Species Database: Gainesville, FL, USA, 2019. Available online: https://nas.er.usgs.gov/queries/factsheet.aspx?SpeciesID=463 (accessed on 2 May 2020).
- Hasan, V.; Tamam, M.B. First record of the invasive Nile Tilapia, Oreochromis niloticus (Linnaeus, 1758) (Perciformes, Cichlidae), on Bawean Island, Indonesia. Check List 2019, 15, 225–227. [Google Scholar] [CrossRef]
- Insani, L.; Hasan, V.; Valen, F.S.; Pratama, F.S.; Widodo, M.S.; Faqih, A.R.; Islamy, R.A.; Mukti, A.T.; Isroni, W. Presence of the invasive nile tilapia Oreochromis niloticus Linnaeus, 1758 (Perciformes, Cichlidae) in the Yamdena Island, Indonesia. Ecol. Environ. Conserv. 2020, 26, 1115–1118. [Google Scholar]
- ISSG. Global Invasive Species Database (GISD) Species Profile Oreochromis mossambicus. 2006. Available online: http://www.iucngisd.org/gisd/species.php?sc=131 (accessed on 28 March 2023).
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume c1979–c2000, pp. 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- He, A.; Luo, Y.; Yang, H.; Liu, L.; Li, S.; Wang, C. Complete mitochondrial DNA sequences of the Nile tilapia (Oreochromis niloticus) and Blue tilapia (Oreochromis aureus): Genome characterization and phylogeny applications. Mol. Biol. Rep. 2011, 38, 2015–2021. [Google Scholar] [CrossRef]
- Aquilino, S.V.; Tango, J.M.; Fontanilla, I.K.; Pagulayan, R.C.; Basiao, Z.U.; Ong, P.S.; Quilang, J.P. DNA barcoding of the ichthyofauna of Taal Lake, Philippines. Mol. Ecol. Resour. 2011, 11, 612–619. [Google Scholar] [CrossRef]
- Mabuchi, K.; Miya, M.; Azuma, Y.; Nishida, M. Independent evolution of the specialized pharyngeal jaw apparatus in cichlid and labrid fishes. BMC Evol. Biol. 2007, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Nwani, C.D.; Becker, S.; Braid, H.E.; Ude, E.F.; Okogwu, O.I.; Hanner, R. DNA barcoding discriminates freshwater fishes from southeastern Nigeria and provides river system-level phylogeographic resolution within some species. Mitochondrial DNA 2011, 22, 43–51. [Google Scholar] [CrossRef] [Green Version]
- ISSG. Global Invasive Species Database (GISD) Species Profile Oreochromis oureus. 2008. Available online: http://www.iucngisd.org/gisd/species.php?sc=1323 (accessed on 2 May 2020).
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Macaranas, J.M.; Taniguchi, N.; Pante, M.J.R.; Capili, J.B.; Pullin, R.S.V. Electrophoretic evidence for extensive hybrid gene introgression into commercial Oreochromis niloticus (L.) stocks in the Philippines. Aquacult. Fish. Manag. 1986, 17, 249–258. [Google Scholar] [CrossRef]
- Cao, J.; Chen, Q.; Lu, M.; Hu, X.; Wang, M. Histology and ultrastructure of the thymus during development in tilapia, Oreochromis niloticus. J. Anat. 2017, 230, 720–733. [Google Scholar] [CrossRef] [Green Version]
- Noble, R.L.; Germany, R.D.; Hall, C.R. Interactions of blue tilapia and largemouth bass in a power plant cooling reservoir. In Proceedings of the Twenty-Ninth Annual Conference, Southeastern Association of Game and Fish Commissioners, St. Louis, MO, USA, 12–15 October 1975; pp. 247–251. [Google Scholar]
- Scoppettone, G.G.; Salgado, J.A.; Nielsen, M.B. Blue tilapia (Oreochromis aureus) predation on fishes in the Muddy River system, Clark County, Nevada. West. N. Am. Nat. 2005, 65, 410–414. [Google Scholar]
- Esselman, P.C. Fish Communities and Conservation of Aquatic Landscapes in Northeastern Mesoamerica; University of Michigan: Ann Arbor, MI, USA, 2009. [Google Scholar]
- ISSG. Global Invasive Species Database (GISD) Species Profile Oreochromis niloticus. 2008. Available online: http://www.iucngisd.org/gisd/species.php?sc=1322 (accessed on 2 May 2020).
- McCrary, J.K.; Murphy, B.R.; Stauffer, J.R.; Hendrix, S.S. Tilapia (Teleostei: Cichlidae) status in Nicaraguan natural waters. Environ. Biol. Fishes. 2007, 78, 107–114. [Google Scholar] [CrossRef]
- Bittencourt, L.S.; Silva, U.R.L.; Silva, L.M.A.; Dias, M.T. Impact of the invasion from Nile tilapia on natives Cichlidae species in tributary of Amazonas River, Brazil. Biota Amaz. 2014, 4, 88–94. [Google Scholar] [CrossRef]
- Marr, S.; Gouws, G.; Avlijas, S.; Khosa, D.; Impson, N.; van der Westhuizen, M.; Weyl, O. Record of Blue tilapia Oreochromis aureus (Steindachner, 1864) in the Eerste River catchment, Western Cape province, South Africa. Afr. J. Aquat. Sci. 2018, 43, 187–193. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphological Comparison | Blue Tilapia (O. aureus) | Nile Tilapia (O. niloticus) | Mozambique Tilapia (O. mossambicus) |
|---|---|---|---|
| Dorsal spines | 15–16 | 16–18 | 15–18 |
| Total dorsal spines and rays | 27–30 | 29–31 | 10–13 |
| Gill rakers | 18–26 | 27–33 | 14–20 |
| Banding on caudal fin | No distinct bands/stripes | Distinct, regular dark stripes | No distinct bands |
| Male breeding coloration | Metallic blue | Red | Black |
| Dorsal fin margin | Vermilion | Dark grey or black | Red |
| Species | GenBank Accession Number | Reference | Sampling Area |
|---|---|---|---|
| Oreochromis aureus | GU370125 | [37] | Africa |
| Oreochromis niloticus | HQ654742 | [38] | Philippines |
| KT307765 | [23] | Philippines | |
| KT307766 | |||
| GU370126 | [37] | Africa | |
| O. niloticus × O. mossambicus | DQ426668 | unpublished | China |
| O. mossambicus × O. niloticus | JQ742044 | unpublished | unknown |
| JQ742045 | |||
| Oreochromis niloticus GIFT | GU477624 | unpublished | unknown |
| Oreochromis sp. | AP009126 | [39] | unknown |
| Tilapia zillii | HM882906 | [40] | Nigeria |
| JF510525 |
| Morphological Comparison | Blue Tilapia (O. aureus) | Nile Tilapia (O. niloticus) |
|---|---|---|
| Dorsal spines | 14–16 (average 15.1 ± 0.8) | 15–17 (average 15.4 ± 0.8) |
| Total dorsal spines and rays | 26–28 (average 27.1 ± 0.9) | 26–29 (average 27.4 ± 0.9) |
| Gill rakers | 23–26 (average 24.0 ± 1.1) | 28–31 (average 29.4 ± 1.0) |
| Banding on caudal fin | No distinct bands/stripes | Distinct, regular dark stripes |
| Dorsal fin margin | Vermilion | Dark grey or black |
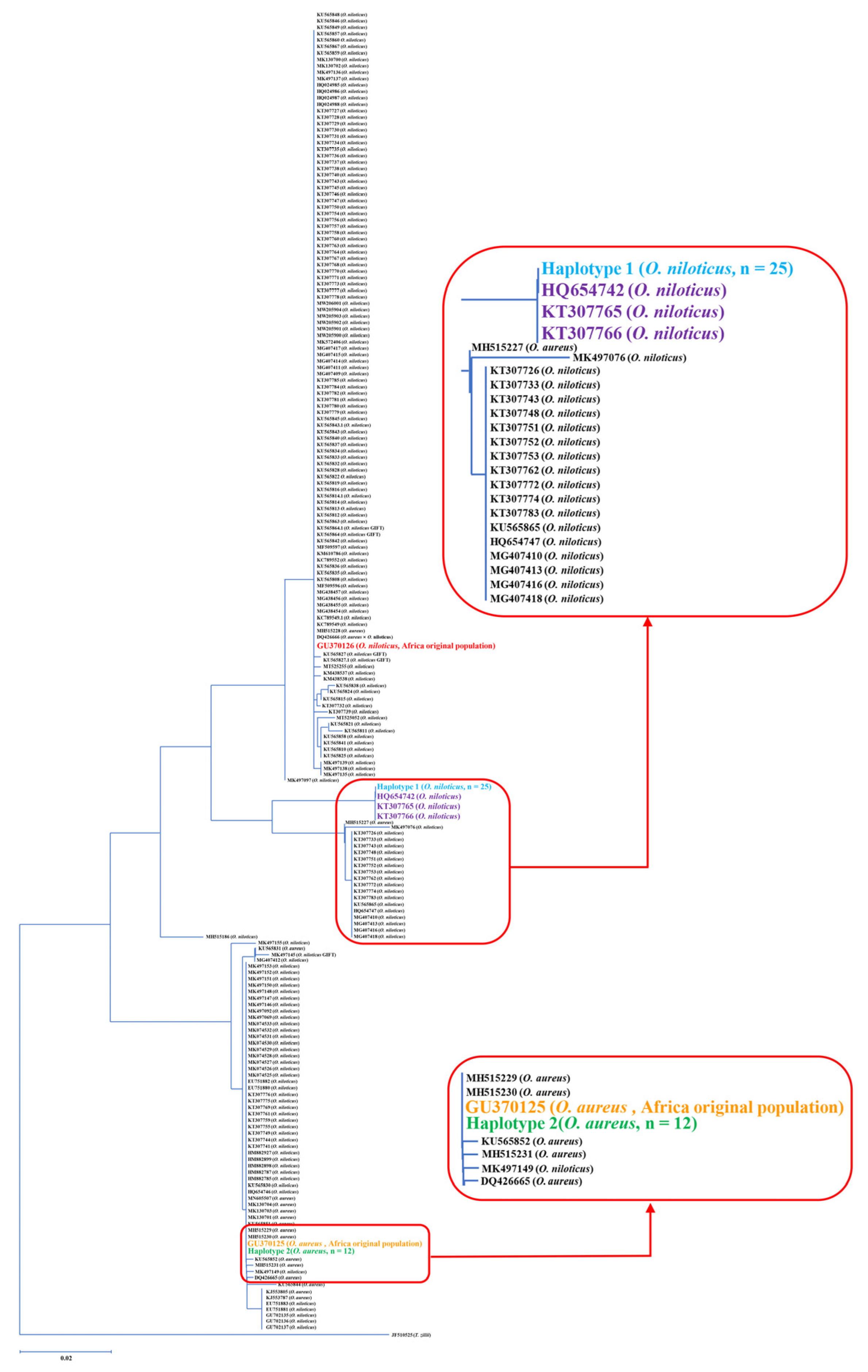
| Species Designation | Morphological Identification | Frequency of Each Haplotype Per Specimen Designation | No. of Samples | Genetic Identification/GenBank Accession Number | |
|---|---|---|---|---|---|
| Haplotype 1 | Haplotype 2 | ||||
| O. aureus | 3, 7, 8, 9, 11, 16, 17, 33, 34, 35, 36, 37 | - | 3, 7, 8, 9, 11, 16, 17, 33, 34, 35, 36, 37 | 12 | O. aureus/GU370125 |
| O. niloticus | 1, 2, 4, 5, 6, 10, 12, 13, 14, 15, 18, 19, 20, 21, 22, 23, 25, 25, 26, 27, 28, 29, 30, 31, 32 | 1, 2, 4, 5, 6, 10, 12, 13, 14, 15, 18, 19, 20, 21, 22, 23, 25, 25, 26, 27, 28, 29, 30, 31, 32 | - | 25 | O. niloticus/HQ654742, KT307765, KT307766 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.H.; Choi, H.-k.; Lee, H.J.; Lee, H.G. On the Species Identification of Two Non-Native Tilapia Species, Including the First Record of a Feral Population of Oreochromis aureus (Steindachner, 1864) in South Korea. Animals 2023, 13, 1351. https://doi.org/10.3390/ani13081351
Wang JH, Choi H-k, Lee HJ, Lee HG. On the Species Identification of Two Non-Native Tilapia Species, Including the First Record of a Feral Population of Oreochromis aureus (Steindachner, 1864) in South Korea. Animals. 2023; 13(8):1351. https://doi.org/10.3390/ani13081351
Chicago/Turabian StyleWang, Ju Hyoun, Hee-kyu Choi, Hyuk Je Lee, and Hwang Goo Lee. 2023. "On the Species Identification of Two Non-Native Tilapia Species, Including the First Record of a Feral Population of Oreochromis aureus (Steindachner, 1864) in South Korea" Animals 13, no. 8: 1351. https://doi.org/10.3390/ani13081351