
Karyotypic and Molecular Analysis of Pterygoplichthys pardalis (Castelnau 1855) from the Lower Amazon River
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Study Area
2.2. Ethical Statements
2.3. Chromosome Preparation and Cytogenetic Analysis
2.4. Molecular Methods
3. Results
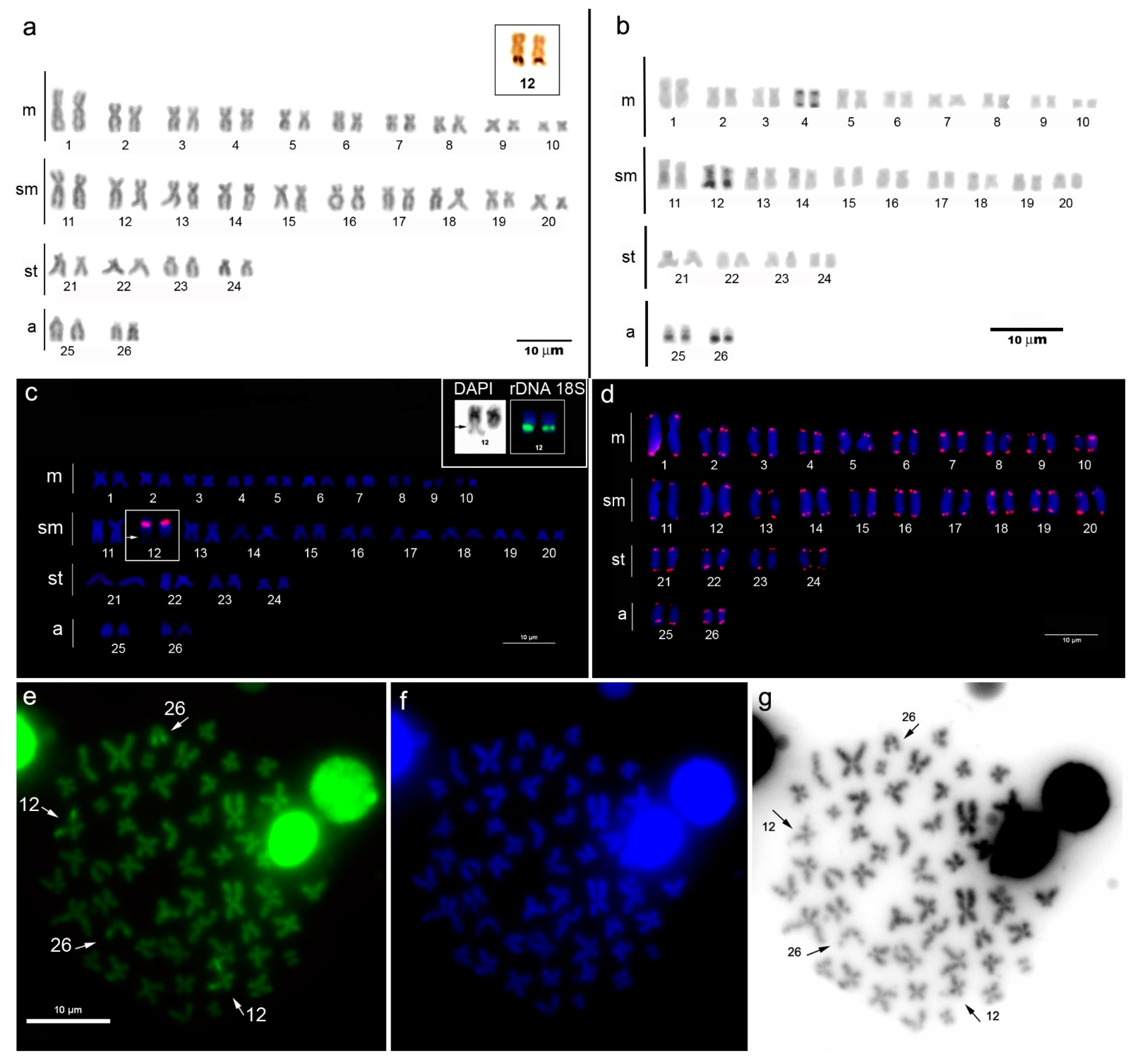
3.1. Cytogenetic Analysis
3.2. Molecular Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fricke, R.; Eschmeyer, W.N.; van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 4 March 2022).
- Lujan, N.K.; Armbruster, J.W.; Lovejoy, N.R.; López-Fernández, H. Multilocus Molecular Phylogeny of the Suckermouth Armored Catfishes (Siluriformes: Loricariidae) with a Focus on Subfamily Hypostominae. Mol. Phylogenetics Evol. 2015, 82, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Reis, R.E.; Britski, H.A.; Britto, M.R.; Buckup, P.A.; Calegari, B.B.; Camelier, P.; Delapieve, M.L.S.; Langeani, F.; Lehmann, P.A.; Lucinda, P.H.F.; et al. Poor Taxonomic Sampling Undermines Nomenclatural Stability: A Reply to Roxo et al. Zootaxa 2019, 4701, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Roxo, F.F.; Ochoa, L.E.; Sabaj, M.H.; Lujan, N.K.; Covain, R.; Silva, G.S.C.; Melo, B.F.; Albert, J.S.; Chang, J.; Foresti, F.; et al. Phylogenomic Reappraisal of the Neotropical Catfish Family Loricariidae (Teleostei: Siluriformes) Using Ultraconserved Elements. Mol. Phylogenetics Evol. 2019, 135, 148–165. [Google Scholar] [CrossRef] [PubMed]
- Jardim de Queiroz, L.; Cardoso, Y.; Jacot-des-Combes, C.; Bahechar, I.A.; Lucena, C.A.; Rapp Py-Daniel, L.; Sarmento Soares, L.M.; Nylinder, S.; Oliveira, C.; Parente, T.E.; et al. Evolutionary Units Delimitation and Continental Multilocus Phylogeny of the Hyperdiverse Catfish Genus Hypostomus. Mol. Phylogenetics Evol. 2020, 145, 106711. [Google Scholar] [CrossRef] [PubMed]
- Anjos, M.S.; Bitencourt, J.A.; Nunes, L.A.; Sarmento-Soares, L.M.; Carvalho, D.C.; Armbruster, J.W.; Affonso, P.R.A.M. Species Delimitation Based on Integrative Approach Suggests Reallocation of Genus in Hypostomini Catfish (Siluriformes, Loricariidae). Hydrobiologia 2019, 847, 563–578. [Google Scholar] [CrossRef]
- Reis, R.E. Check List of the Freshwater Fishes of South and Central America; Edipucrs: Porto Alegre, Brazil, 2003. [Google Scholar]
- Orfinger, A.B.; Douglas Goodding, D. The Global Invasion of the Suckermouth Armored Catfish Genus Pterygoplichthys (Siluriformes: Loricariidae): Annotated List of Species, Distributional Summary, and Assessment of Impacts. Zool Stud. 2018, 57, e7. [Google Scholar] [CrossRef]
- Page, L.M.; Robins, R.H. Identification of Sailfin Catfishes (Teleostei: Loricariidae) in Southeastern Asia. Raffles Bull. Zool. 2006, 54, 455–457. [Google Scholar]
- Chavez, J.M.; De La Paz, R.M.; Manohar, S.K.; Pagulayan, R.C.; Vi, J.R.C. New Philippine Record of South American Sailfin Catfishes (Pisces: Loricariidae). Zootaxa 2006, 1109, 57. [Google Scholar] [CrossRef]
- Lozano-Vilano, M.; García-Ramirez, M.; Mendoza, R.; Koleff, P. Peces Invasores En El Noreste de México. In Especies Acuáticas Invasoras en México; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Tlalpan, Mexico, 2014; pp. 401–412. [Google Scholar]
- Ruffino, M.; Soares, E.; Silva, C.; Barthem, R.; Batista, V.; Estupiñan, G.; Pinto, W. Estatística Pesqueira Do Amazonas e Pará; IBAMA: Manaus, Brazil, 2006. [Google Scholar]
- Moroni, F.T.; Moroni, R.B.; Mayag, B.; de Jesus, R.S.; Lessi, E. Limitations in Decision Context for Selection of Amazonian Armoured Catfish Acari-Bod (Pterygoplichthys Pardalis) as Candidate Species for Aquaculture. Int. J. Fish. Aquac. 2015, 7, 142–150. [Google Scholar]
- Ghigliotti, L.; Mazzei, F.; Ozouf-Costaz, C.; Christiansen, J.S.; Fevolden, S.-E.; Pisano, E. First Cytogenetic Characterization of the Sub-Arctic Marine Fish Mallotus villosus (Müller, 1776), Osmeriformes, Osmeridae. Genet. Mol. Biol. 2008, 31 (Suppl. S1), 180–187. [Google Scholar] [CrossRef]
- Vitorino, C.A.; Oliveira, R.C.C.; Margarido, V.P.; Venere, P.C. Genetic Diversity of Arapaima gigas (Schinz, 1822) (Osteoglossiformes: Arapaimidae) in the Araguaia-Tocantins Basin Estimated by ISSR Marker. Neotrop. Ichthyol. 2015, 13, 557–568. [Google Scholar] [CrossRef]
- Hilsdorf, A.R.; Marques, D. Genética e Conservação de Estoques Pesqueiros de Águas Continentais No Brasil: Situação Atual e Perspectivas; Embrapa: Brasilia, Brazil, 2006. [Google Scholar]
- Majtánová, Z.; Moy, K.G.; Unmack, P.J.; Ráb, P.; Ezaz, T. Characterization of the Karyotype and Accumulation of Repetitive Sequences in Australian Darling Hardyhead Craterocephalus amniculus (Atheriniformes, Teleostei). PeerJ 2019, 7, e7347. [Google Scholar] [CrossRef] [PubMed]
- Arai, K. Genetic Improvement of Aquaculture Finfish Species by Chromosome Manipulation Techniques in Japan. Reprod. Biotechnol. Finfish Aquac. 2001, 205–228. [Google Scholar] [CrossRef]
- Bertolini, R.M.; Lopez, L.S.; do Nascimento, N.F.; Arashiro, D.R.; de Siqueira Silva, D.H.; dos Santos, S.C.A.; Senhorini, J.A.; Yasui, G.S. Strategies for Aquaculture and Conservation of Neotropical Catfishes Based on the Production of Triploid Pimelodus maculatus. Aquac. Int. 2019, 28, 127–137. [Google Scholar] [CrossRef]
- Colihueque, N.; Corrales, O.; Parraguez, M. Karyotype and Nuclear DNA Content of Trichomycterus areolatus (Siluriformes, Trichomycteridae). Genet. Mol. Biol. 2006, 29, 278–282. [Google Scholar] [CrossRef]
- Porto-Foresti, F.; Hashimoto, D.T.; Alves, A.L.; Almeida, R.B.C.; Senhorini, J.A.; Bortolozzi, J.; Foresti, F. Cytogenetic Markers as Diagnoses in the Identification of the Hybrid between Piauçu (Leporinus macrocephalus) and Piapara (Leporinus elongatus). Genet. Mol. Biol. 2008, 31 (Suppl. S1), 195–202. [Google Scholar] [CrossRef]
- Artoni, R.F.; Bertollo, L.A.C. Cytogenetic Studies on Hypostominae (Pisces, Siluriformes, Loricariidae). Considerations on Karyotype Evolution in the Genus Hypostomus. Caryologia 1996, 49, 81–90. [Google Scholar] [CrossRef]
- Bueno, V.; Konerat, J.T.; Zawadzki, C.H.; Venere, P.C.; Blanco, D.R.; Margarido, V.P. Divergent Chromosome Evolution in Hypostominae Tribes (Siluriformes: Loricariidae): Correlation of Chromosomal Data with Morphological and Molecular Phylogenies. Zebrafish 2018, 15, 492–503. [Google Scholar] [CrossRef]
- Alves, A.L.; Oliveira, C.; Nirchio, M.; Granado, Á.; Foresti, F. Karyotypic Relationships among the Tribes of Hypostominae (Siluriformes: Loricariidae) with Description of XO Sex Chromosome System in a Neotropical Fish Species. Genetica 2006, 128, 1–9. [Google Scholar] [CrossRef]
- Fernandes, C.A.; Alves, D.S.; Guterres, Z.d.R.; Martins-Santos, I.C. Cytogenetic Analysis of Two Locariid Species (Teleostei, Siluriformes) from Iguatemi River (Parana River Drainage) in Brazil. Comp. Cytogenet. 2015, 9, 67–78. [Google Scholar] [CrossRef]
- Artoni, R.F.; Molina, W.F.; Bertollo, L.A.C.; Galetti Junior, P.M. Heterochromatin analysis in the fish species Liposarcus anisitsi (siluriformes) and Leporinus elongatus (characiformes). Genet. Mol. Biol. 1999, 22, 39–44. [Google Scholar] [CrossRef]
- Alves, A.L.; de Borba, R.S.; Pozzobon, A.P.B.; Oliveira, C.; Nirchio, M.; Granado, A.; Foresti, F. Localization of 18S Ribosomal Genes in Suckermouth Armoured Catfishes Loricariidae (Teleostei, Siluriformes) with Discussion on the Ag-NOR Evolution. Comp. Cytogenet. 2012, 6, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.R.d.; Souza, I.L.; Venere, P.C. Karyotype Description of Three Species of Loricariidae (Siluriformes) and Occurrence of the ZZ/ZW Sexual System in Hemiancistrus spilomma Cardoso & Lucinda, 2003. Neotrop. Ichthyol. 2006, 4, 93–97. [Google Scholar] [CrossRef]
- Araújo da Silva, F.; Feldberg, E.; Moura Carvalho, N.D.; Hernández Rangel, S.M.; Schneider, C.H.; Carvalho-Zilse, G.A.; Fonsêca da Silva, V.; Gross, M.C. Effects of Environmental Pollution on the RDNAomics of Amazonian Fish. Environ. Pollut. 2019, 252, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Toledo, L.F.A.; Foresti, F.; Toledo, S.A. Supernumerary Chromosomes, Robertsonian Rearrangement and Multiple NORs In Corydoras aeneus (Pisces, Siluriformes, Callichthyidae). Caryologia 1988, 41, 227–236. [Google Scholar] [CrossRef]
- Howell, W.M.; Black, D.A. Controlled Silver-Staining of Nucleolus Organizer Regions with a Protective Colloidal Developer: A 1-Step Method. Experientia 1980, 36, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Sumner, A.T. A Simple Technique for Demonstrating Centromeric Heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Lui, R.; Blanco, D.; Moreira-Filho, O.; Margarido, V. Propidium Iodide for Making Heterochromatin More Evident in the C-Banding Technique. Biotech. Histochem. 2012, 87, 433–438. [Google Scholar] [CrossRef]
- Donlon, T.A.; Magenis, R.E. Methyl Green Is a Substitute for Distamycin A in the Formation of Distamycin A/DAPI C-Bands. Hum. Genet. 1983, 65, 144–146. [Google Scholar] [CrossRef]
- Schweizer, D. Reverse Fluorescent Chromosome Banding with Chromomycin and DAPI. Chromosoma 1976, 58, 307–324. [Google Scholar] [CrossRef]
- Pinkel, D.; Straume, T.; Gray, J.W. Cytogenetic Analysis Using Quantitative, High-Sensitivity, Fluorescence Hybridization. Proc. Natl. Acad. Sci. USA 1986, 83, 2934–2938. [Google Scholar] [CrossRef] [PubMed]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved Telomere Detection Using a Telomere Repeat Probe (TTAGGG)n Generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef] [PubMed]
- Komiya, H.; Takemura, S. Nucleotide Sequence of 5S Ribosomal RNA from Rainbow Trout (Salmo gairdnerii) Liver1. J. Biochem. 1979, 86, 1067–1080. [Google Scholar] [CrossRef]
- Guerra, M. Citogenética Molecular: Protocolos Comentados. In Sociedade Brasileira de Genética, Ribeirão Preto; Sociedade Brasileira de Genética: Ribeirao Preto, Brazil, 2012. [Google Scholar]
- Aljanabi, S.; Martinez, I. Universal and Rapid Salt-Extraction of High-Quality Genomic DNA for PCR- Based Techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA Barcoding Australia’s Fish Species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Dunn, I.S.; Blattner, F.R. Charons 36 to 40: Multi Enzyme, High Capacity, Recombination Deficient Replacement Vectors with Polylinkers and Polystuffers. Nucleic Acids Res. 1987, 15, 2677–2698. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London. UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1. 3.1; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2010. [Google Scholar]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A Software for Comprehensive Analysis of DNA Polymorphism Data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Nirchio, M.; Rossi, A.R.; Foresti, F.; Oliveira, C. Chromosome Evolution in Fishes: A New Challenging Proposal from Neotropical Species. Neotrop. Ichthyol. 2014, 12, 761–770. [Google Scholar] [CrossRef]
- Neves, A.M.; Ruffino, M.L. Aspectos reprodutivos do acarí-bodó Liposarcus pardalis (Pisces, Siluriformes, Loricariidae) (Castelnau, 1855) do Médio Amazonas. Bol. Do Mus. Para. Emílio Goeldi Série. Zool. 1998, 14, 77–94. [Google Scholar]
- Pety, A.M.; Cardoso, A.L.; Nagamachi, C.Y.; Pieczarka, J.C.; de Sousa, L.M.; Noronha, R.C.R. In Situ Localization of Ribosomal Sites in Peckoltia and Ancistomus (Loricariidae: Hypostominae) from the Amazon Basin. Zebrafish 2018, 15, 263–269. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, L.C.; Ribeiro, M.O.; Costa, G. de M.; Zawadzki, C.H.; Prizon-Nakajima, A.C.; Borin-Carvalho, L.A.; Martins-Santos, I.C.; Portela-Castro, A.L. de B. Cytogenetic Characterization of Hypostomus soniae Hollanda-Carvalho & Weber, 2004 from the Teles Pires River, Southern Amazon Basin: Evidence of an Early Stage of an XX/XY Sex Chromosome System. Comp. Cytogenet. 2019, 13, 411–422. [Google Scholar] [CrossRef]
- Traldi, J.B.; Vicari, M.R.; Blanco, D.R.; Martinez, J.d.F.; Artoni, R.F.; Moreira-Filho, O. First Karyotype Description of Hypostomus iheringii (Regan, 1908): A Case of Heterochromatic Polymorphism. Comp. Cytogenet. 2012, 6, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Gornung, E. Twenty Years of Physical Mapping of Major Ribosomal RNA Genes across the Teleosts: A Review of Research. Cytogenet. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Gosztonyi, A.E. A Cytogenetic Study of Diplotnystes mesembrinus (Teleostei, Siluriformes, Diplomystidae) with a Discussion of Chromosome Evolution in Siluriforms. Caryologia 2000, 53, 31–37. [Google Scholar] [CrossRef]
- Ziemniczak, K.; Barros, A.V.; Rosa, K.O.; Nogaroto, V.; Almeida, M.C.; Cestari, M.M.; Moreira-Filho, O.; Artoni, R.F.; Vicari, M.R. Comparative Cytogenetics of Loricariidae (Actinopterygii: Siluriformes): Emphasis in Neoplecostominae and Hypoptopomatinae. Ital. J. Zool. 2012, 79, 492–501. [Google Scholar] [CrossRef]
- Artoni, R.F.; Carlos Bertollo, L.A. Trends in the Karyotype Evolution of Loricariidae Fish (Siluriformes). Hereditas 2004, 134, 201–210. [Google Scholar] [CrossRef]
- Ardura, A.; Linde, A.R.; Moreira, J.C.; Garcia-Vazquez, E. DNA Barcoding for Conservation and Management of Amazonian Commercial Fish. Biol. Conserv. 2010, 143, 1438–1443. [Google Scholar] [CrossRef]
- Vargas-Rivas, A.G.; Barba-Macias, E.; Sánchez, A.J.; Castellanos-Morales, G. Lack of MtDNA Genetic Diversity despite Phenotypic Variation and Environmental Heterogeneity in the Exotic Suckermouth Armored Catfish (Pterygoplichthys pardalis). Biol. Invasions 2022, 25, 1035–1056. [Google Scholar] [CrossRef]
- Jumawan, J.C.; Vallejo, B.M.; Herrera, A.A.; Buerano, C.C.; Fontanilla, I.K.C. DNA Barcodes of the Suckermouth Sailfin Catfish Pterygoplichthys (Siluriformes: Loricariidae) in the Marikina River System, Philippines: Molecular Perspective of an Invasive Alien Fish Species. Philipp. Sci. Lett. 2011, 4, 103–113. [Google Scholar]
- Yu, S.C.S.; Quilang, J.P. Molecular Phylogeny of Catfishes (Teleostei: Siluriformes) in the Philippines Using the Mitochondrial Genes COI, Cyt b, 16S RRNA, and the Nuclear Genes Rag1 and Rag2. Philipp. J. Sci. 2014, 143, 187–198. [Google Scholar]
- Yonekura, R.; Kawamura, K.; Uchii, K.A. Peculiar Relationship between Genetic Diversity and Adaptability in Invasive Exotic Species: Bluegill Sunfish as a Model Species. Ecol. Res. 2007, 22, 911–919. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The Population Biology of Invasive Species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef]
- Wu, L.W.; Liu, C.C.; Lin, S.-M. Identification of Exotic Sailfin Catfish Species (Pterygoplichthys, Loricariidae) in Taiwan Based on Morphology and MtDNA Sequences. Zool. Stud. 2011, 50, 235–246. [Google Scholar]
- Tomé-Souza, M.J.F.; Raseira, M.B.; Ruffino, M.L.; Silva, C.O.; Batista, V.S.; Barthem, R.B.; Amaral, E.S. Estatística Pesqueira do Amazonas e Pará—2004; Ibama/ProVárzea: Manaus, Brazil, 2007. [Google Scholar]
- Castello, L.; McGrath, D.G.; Arantes, C.C.; Almeida, O.T. Accounting for Heterogeneity in Small-Scale Fisheries Management: The Amazon Case. Mar. Policy 2013, 38, 557–565. [Google Scholar] [CrossRef]
- Castello, L.; McGrath, D.G.; Beck, P.S.A. Resource Sustainability in Small-Scale Fisheries in the Lower Amazon Floodplains. Fish. Res. 2011, 110, 356–364. [Google Scholar] [CrossRef]
- Marinho Lima, E.M.; Brasil Santos, P.R.; Porto Braga, T.M.; Gibbs McGrath, D. A pesca de acari (Pterygoplichthys pardalis) na várzea do Baixo Amazonas, Pará, Brasil: Aspectos estruturais e socioeconômicos. Gaia Sci. 2019, 13, 1268–1981. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Locality | KF (m + sm + st + a) | BC | NOR | CMA3 | 18S | 5S | Rex1 | Rex3 | Rex6 | Authors |
|---|---|---|---|---|---|---|---|---|---|---|---|
| P. anisitsi | rio Tietê | 28 + 12 + 8 + 4 | - | 16 | - | - | - | - | - | - | [24] |
| P. anisitsi | rio Miranda | 8 + 14 + 14 + 16 | - | 5 | - | - | - | - | - | - | [24] |
| P. anisitsi | rio Iguatemi | 14 + 26 + 8 + 4 | + | 9 | 9, 10 | - | - | - | - | - | [25] |
| P. anisitsi | rio Preto | 16 + 24 + 8 + 4 | + | 14 | - | - | - | - | - | - | [26] |
| P. ambrosettii | rio Paraná | 16 + 24 + 8 + 4 | - | - | - | 14 | 14 | - | - | - | [23] |
| P. multiradiatus | rio Orinoco | 22 + 18 + 12 + 0 | - | 10 | - | 10 | - | - | - | - | [24,27] |
| P. gibbiceps | rio Orinoco | 20 + 24 + 8 + 0 | - | 21 | - | - | - | - | - | - | [24] |
| P. joselimaianus | rio Araguaia | 28 + 16 + 8 + 0 | - | 9 | - | - | - | - | - | - | [28] |
| P. pardalis | rio Amazonas | 18 + 18 + 8 + 8 | - | 11 | - | 11 | 11 | + | + | + | [29] |
| 1 | 2 | 3 | 4 | 5 | 6 | ||
|---|---|---|---|---|---|---|---|
| 1 | Hypostomus cochliodon | ||||||
| 2 | Pterygoplichthys etentaculatus | 0.078 | |||||
| 3 | P. pardalis (Asia) | 0.086 | 0.031 | ||||
| 4 | P. pardalis (Colombia) | 0.084 | 0.030 | 0.000 | |||
| 5 | P. pardalis (Manaus, Brazil) | 0.089 | 0.030 | 0.000 | 0.000 | ||
| 6 | P. pardalis (Santarém, Brazil) | 0.086 | 0.031 | 0.000 | 0.000 | 0.000 | |
| 7 | P. zuliaensis | 0.127 | 0.068 | 0.075 | 0.071 | 0.085 | 0.075 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos Guimarães, A.; Maciel, L.A.M.; de Souza, M.F.B.; Rodrigues, L.R.R. Karyotypic and Molecular Analysis of Pterygoplichthys pardalis (Castelnau 1855) from the Lower Amazon River. Animals 2023, 13, 1533. https://doi.org/10.3390/ani13091533
dos Santos Guimarães A, Maciel LAM, de Souza MFB, Rodrigues LRR. Karyotypic and Molecular Analysis of Pterygoplichthys pardalis (Castelnau 1855) from the Lower Amazon River. Animals. 2023; 13(9):1533. https://doi.org/10.3390/ani13091533
Chicago/Turabian Styledos Santos Guimarães, Alcimara, Luan Aércio Melo Maciel, Mendelshon Fujiie Belém de Souza, and Luís Reginaldo Ribeiro Rodrigues. 2023. "Karyotypic and Molecular Analysis of Pterygoplichthys pardalis (Castelnau 1855) from the Lower Amazon River" Animals 13, no. 9: 1533. https://doi.org/10.3390/ani13091533