
A Comparative Study of the Fatty Acids and Monosaccharides of Wild and Cultivated Ulva sp.

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
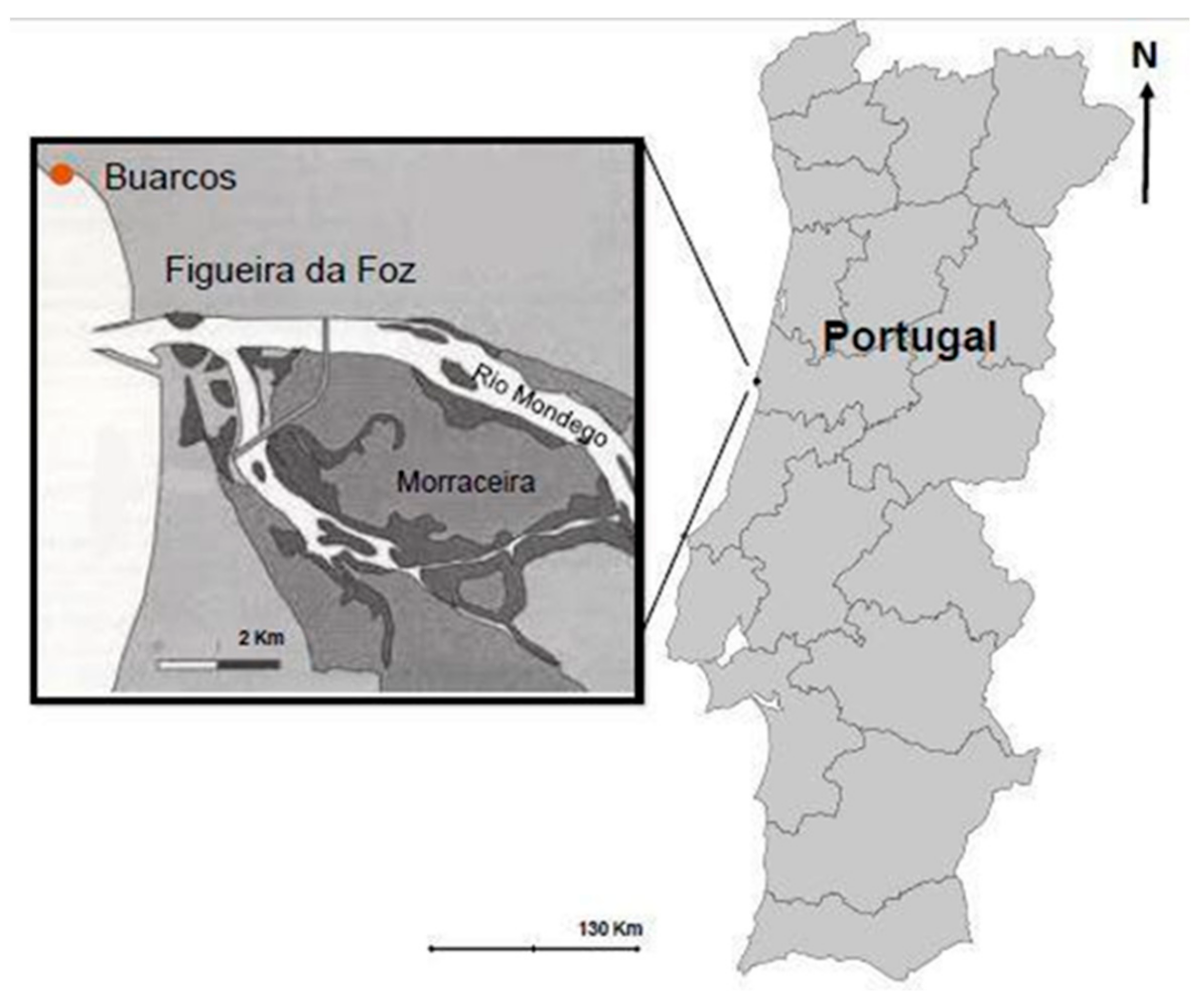
2.1. Seaweed Collection
2.2. Seaweed Cultivation Method
2.2.1. Indoor Cultivation (Aquaria)
2.2.2. Inshore Cultivation (Tank)
2.2.3. Quantitative Analysis of the Seaweed Growth
2.3. Samples Preparation for Biochemical Analysis
2.3.1. Fatty Acids Analysis by GC/MS
2.3.2. Carbohydrate Analysis by GC-FID
2.4. Data Analyses
3. Results
3.1. Ulva sp. Cultivation Growth
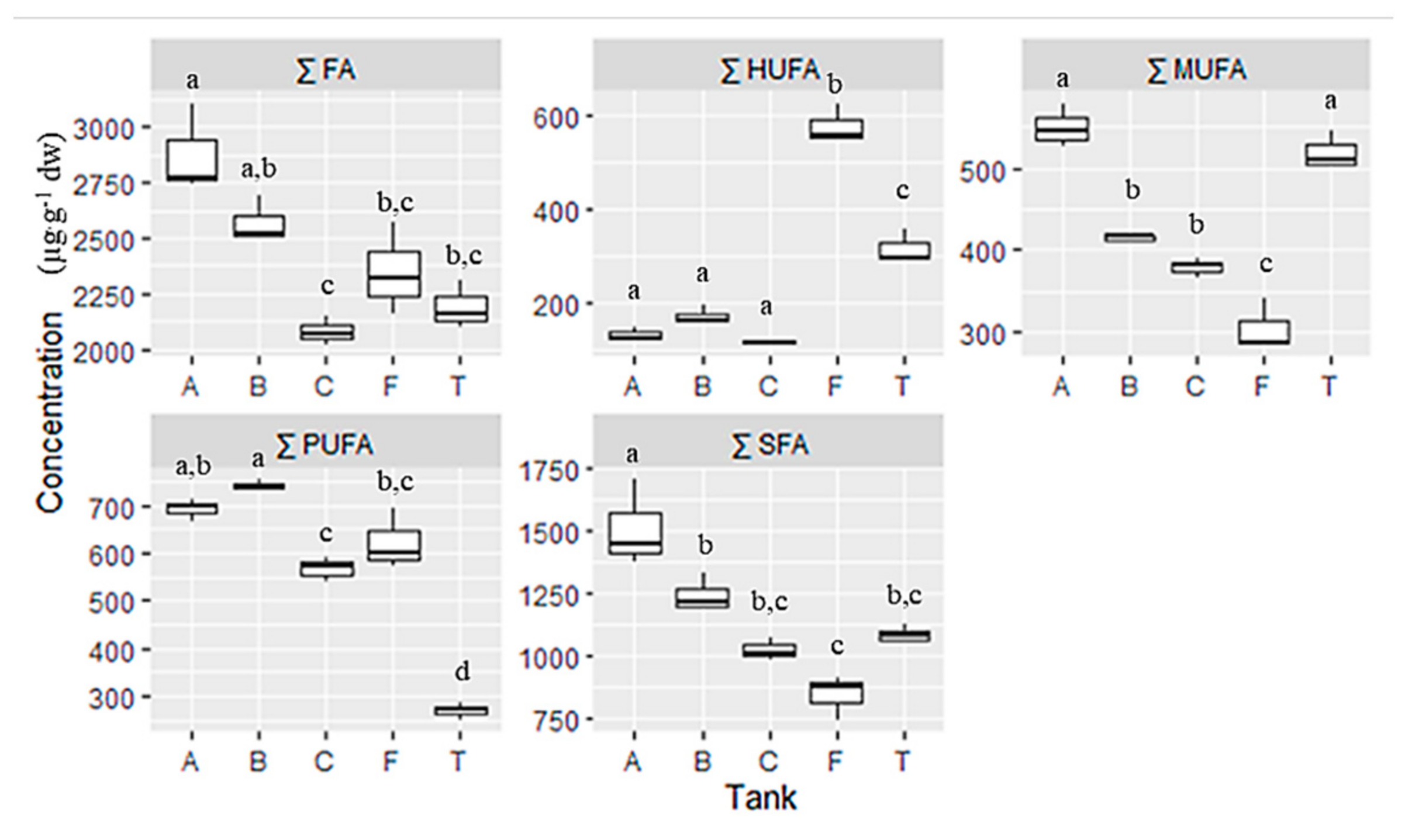
3.2. Fatty Acid Characterization
3.3. Carbohydrate Characterization
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pereira, L. As Algas Marinhas e Respectivas Utilidades; Coimbra. 2010. Available online: https://www.researchgate.net/publication/341056597_As_Algas_Marinhas_e_Respectivas_Utilidades (accessed on 3 February 2022).
- Leandro, A.; Pacheco, D.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. Seaweed’s bioactive candidate compounds to food industry and global food security. Life 2020, 10, 140. [Google Scholar] [CrossRef]
- García-Poza, S.; Leandro, A.; Cotas, C.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. The evolution road of seaweed aquaculture: Cultivation technologies and the industry 4.0. Int. J. Environ. Res. Public Health 2020, 17, 6528. [Google Scholar] [CrossRef] [PubMed]
- GBIF Secretariat GBIF Backbone Taxonomy. Available online: https://www.gbif.org/dataset/d7dddbf4-2cf0-4f39-9b2a-bb099caae36c (accessed on 3 January 2022).
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eladel, H.; Moustafa, Y.T.A.; Eladel, H.M. Amino acids profile of Ulva sp. macroalgae collected from Alexandria coast in Egypt: A potential food resource. Bot. Microbiol. Res. 2015, 1, 015–022. [Google Scholar]
- Kraft, L.G.K.; Kraft, G.T.; Waller, R.F. Investigations into southern australian Ulva (Ulvophyceae, Chlorophyta) Taxonomy and molecular phylogeny indicate both cosmopolitanism and endemic cryptic species1. J. Phycol. 2010, 46, 1257–1277. [Google Scholar] [CrossRef]
- Ben-Ari, T.; Neori, A.; Ben-Ezra, D.; Shauli, L.; Odintsov, V.; Shpigel, M. Management of Ulva lactuca as a biofilter of mariculture effluents in IMTA system. Aquaculture 2014, 434, 493–498. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, J.; Gao, S.; Huo, Y.; Cui, J.; Shen, H.; Liu, G.; He, P. Annual patterns of macroalgal blooms in the Yellow Sea during 2007–2017. PLoS ONE 2019, 14, e0210460. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Huo, Y.; Wu, H.; Yu, K.; Kim, J.K.; Yarish, C.; Qin, Y.; Liu, C.; Xu, R.; He, P. The origin of the Ulva macroalgal blooms in the Yellow Sea in 2013. Mar. Pollut. Bull. 2014, 89, 276–283. [Google Scholar] [CrossRef]
- Liu, J.; Zhuang, M.; Zhao, L.; Liu, Y.; Wen, Q.; Fu, M.; Yu, K.; Zhang, J.; He, P. Taxonomy and Genetic Diversity of Amphipods Living on Ulva lactuca L. from Gouqi Coast, China1. Pac. Sci. 2020, 74, 137–146. [Google Scholar] [CrossRef]
- Liu, J.; Xia, J.; Zhuang, M.; Zhang, J.; Yu, K.; Zhao, S.; Sun, Y.; Tong, Y.; Xia, L.; Qin, Y.; et al. Controlling the source of green tides in the Yellow Sea: NaClO treatment of Ulva attached on Pyropia aquaculture rafts. Aquaculture 2021, 535, 736378. [Google Scholar] [CrossRef]
- Liu, J.; Li, C.; Xia, J.; Sun, Y.; Tong, Y.; Zhang, J.; Zhao, S.; Zhuang, M.; He, P. Epizoic Ulva attached to intertidal animals in the Subei intertidal zone are not the additional source of the famed Yellow Sea green tides. J. Sea Res. 2021, 174, 102065. [Google Scholar] [CrossRef]
- Liu, J.; Tong, Y.; Xia, J.; Sun, Y.; Zhao, X.; Sun, J.; Zhao, S.; Zhuang, M.; Zhang, J.; He, P. Ulva macroalgae within local aquaculture ponds along the estuary of Dagu River, Jiaozhou Bay, Qingdao. Mar. Pollut. Bull. 2022, 174, 113243. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Fattah, A.F.; Edrees, M. Seasonal changes in the constituents of Ulva lactuca. Phytochemistry 1973, 12, 481–485. [Google Scholar] [CrossRef]
- Ortiz, J.; Romero, N.; Robert, P.; Araya, J.; Lopez-Hernández, J.; Bozzo, C.; Navarrete, E.; Osorio, A.; Rios, A. Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea antarctica. Food Chem. 2006, 99, 98–104. [Google Scholar] [CrossRef]
- Gonçalves, A.M.M.; Marques, J.C.; Gonçalves, F. Fatty acids’ profiles of aquatic organisms: Revealing the impacts of environmental and anthropogenic stressors. In Fatty Acids; Catala, A., Ed.; IntechOpen: London, UK, 2017. [Google Scholar]
- De Alencar, D.B.; Diniz, J.C.; Rocha, S.A.S.; Pires-Cavalcante, K.M.S.; De Lima, R.L.; De Sousa, K.C.; Freitas, J.O.; Bezerra, R.M.; Baracho, B.M.; Sampaio, A.H.; et al. Fatty acid composition from the marine red algae Pterocladiella capillacea (S. G. gmelin) santelices & hommersand 1997 and Osmundaria obtusiloba (C. agardh) R. E. norris 1991 and its antioxidant activity. An. Acad. Bras. Cienc. 2018, 90, 449–459. [Google Scholar] [CrossRef]
- Kendel, M.; Wielgosz-Collin, G.; Bertrand, S.; Roussakis, C.; Bourgougnon, N.; Bedoux, G. Lipid composition, fatty acids and sterols in the seaweeds Ulva armoricana, and Solieria chordalis from Brittany (France): An analysis from nutritional, chemotaxonomic, and antiproliferative activity perspectives. Mar. Drugs 2015, 13, 5606–5628. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Yaich, H.; Garna, H.; Besbes, S.; Paquot, M.; Blecker, C.; Attia, H. Chemical composition and functional properties of Ulva lactuca seaweed collected in Tunisia. Food Chem. 2011, 128, 895–901. [Google Scholar] [CrossRef]
- Santos, S.A.O.; Vilela, C.; Freire, C.S.R.; Abreu, M.H.; Rocha, S.M.; Silvestre, A.J.D. Chlorophyta and Rhodophyta macroalgae: A source of health promoting phytochemicals. Food Chem. 2015, 183, 122–128. [Google Scholar] [CrossRef]
- Peña-Rodríguez, A.; Mawhinney, T.P.; Ricque-Marie, D.; Cruz-Suárez, L.E. Chemical composition of cultivated seaweed Ulva clathrata (Roth) C. Agardh. Food Chem. 2011, 129, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.; Guo, Z.; Yang, Y.; Geng, Z.; Tang, L.; Zhao, M.; Qiu, Y.; Chen, Y.; He, P. Inhibition of hydrogen peroxide induced injuring on human skin fibroblast by Ulva prolifera polysaccharide. Int. J. Biol. Macromol. 2016, 91, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Almeida, V.L. The marine macroalgae of the genus Ulva: Chemistry, biological activities and potential applications. Oceanogr. Open Access 2013, 1, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-rodríguez, J.A.; Amaya-guerra, C.A. Ulva genus as alternative crop: Nutritional and functional properties. In Alternative Crops and Cropping Systems; Konvalina, P., Ed.; IntechOpen: London, UK, 2015; pp. 29–44. Available online: https://www.intechopen.com/chapters/50340 (accessed on 3 January 2022).
- Mangott, A.; Nappi, J.; Paoli, A.D.; Goncalves, P.; Hua, K.; Domingos, J.A.; De Nys, R.; Thomas, T. Ulva lactuca as a functional ingredient and water bioremediator positively influences the hepatopancreas and water microbiota in the rearing of Litopenaeus vannamei. Algal Res. 2020, 51, 102040. [Google Scholar] [CrossRef]
- Steinhagen, S.; Enge, S.; Larsson, K.; Olsson, J.; Nylund, G.M.; Albers, E.; Pavia, H.; Undeland, I.; Toth, G.B. Sustainable large-scale aquaculture of the northern hemisphere sea lettuce, Ulva fenestrata, in an off-shore seafarm. J. Mar. Sci. Eng. 2021, 9, 615. [Google Scholar] [CrossRef]
- Nesterov, V.N.; Rozentsvet, O.A.; Bogdanova, E.S. Influence of abiotic factors on the content of fatty acids of Ulva Intestinalis. Contemp. Probl. Ecol. 2013, 6, 441–447. [Google Scholar] [CrossRef]
- Pereira, L. Ulva lactuca Linnaeus. Available online: http://macoi.uc.pt/spec_list_detail.php?spec_id=171&searchSpecies=ulva+lactuca+linnaeus (accessed on 17 May 2021).
- Vandermeulen, H.; Gordin, H. Ammonium uptake using Ulva (Chlorophyta) in intensive fishpond systems: Mass culture and treatment of effluent. J. Appl. Phycol. 1990, 2, 363–374. [Google Scholar] [CrossRef]
- Araujo, G.S.; Cotas, J.; Morais, T.; Leandro, A.; García-Poza, S.; Gonçalves, A.M.M.; Pereira, L. Calliblepharis jubata cultivation potential—A comparative study between controlled and semi-controlled aquaculture. Appl. Sci. 2020, 10, 7553. [Google Scholar] [CrossRef]
- Pérez-Mayorga, D.M.; Ladah, L.B.; Zertuche-González, J.A.; Leichter, J.J.; Filonov, A.E.; Lavín, M.F. Nitrogen uptake and growth by the opportunistic macroalga Ulva lactuca (Linnaeus) during the internal tide. J. Exp. Mar. Bio. Ecol. 2011, 406, 108–115. [Google Scholar] [CrossRef]
- Gonçalves, A.M.M.; Pardal, M.Â.; Marques, S.C.; Mendes, S.; Fernández-Gómez, M.J.; Galindo-Villardón, M.P.; Azeiteiro, U.M. Responses of copepoda life-history stages to climatic variability in a southern-European temperate estuary. Zool. Stud. 2012, 51, 321–335. [Google Scholar]
- Coimbra, M.A.; Waldron, K.W.; Selvendran, R.R. Isolation and characterisation of cell wall polymers from olive pulp (Olea europaea L.). Carbohydr. Res. 1994, 252, 245–262. [Google Scholar] [CrossRef]
- Human, L.R.D.; Adams, J.B.; Allanson, B.R. Insights into the cause of an Ulva lactuca Linnaeus bloom in the Knysna Estuary. S. Afr. J. Bot. 2016, 107, 55–62. [Google Scholar] [CrossRef]
- Tang, Y.Z.; Gobler, C.J. The green macroalga, Ulva lactuca, inhibits the growth of seven common harmful algal bloom species via allelopathy. Harmful Algae 2011, 10, 480–488. [Google Scholar] [CrossRef]
- Gupta, V.; Kushwaha, H.R. Metabolic regulatory oscillations in intertidal green seaweed Ulva lactuca against tidal cycles. Sci. Rep. 2017, 7, 16430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Msuya, F.E.; Neori, A. Effect of water aeration and nutrient load level on biomass yield, N uptake and protein content of the seaweed Ulva lactuca cultured in seawater tanks. J. Appl. Phycol. 2008, 20, 1021–1031. [Google Scholar] [CrossRef]
- Queirós, A.S.; Circuncisão, A.R.; Pereira, E.; Válega, M.; Abreu, M.H.; Silva, A.M.S.; Cardoso, S.M. Valuable nutrients from Ulva rigida: Modulation by seasonal and cultivation factors. Appl. Sci. 2021, 11, 6137. [Google Scholar] [CrossRef]
- Califano, G.; Kwantes, M.; Abreu, M.H.; Costa, R.; Wichard, T. Cultivating the macroalgal holobiont: Effects of integrated multi-trophic aquaculture on the microbiome of Ulva rigida (Chlorophyta). Front. Mar. Sci. 2020, 7, 52. [Google Scholar] [CrossRef]
- Wichard, T. Exploring bacteria-induced growth and morphogenesis in the green macroalga order Ulvales (Chlorophyta). Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Alsufyani, T.; Califano, G.; Deicke, M.; Grueneberg, J.; Weiss, A.; Engelen, A.H.; Kwantes, M.; Mohr, J.F.; Ulrich, J.F.; Wichard, T. Macroalgal–bacterial interactions: Identification and role of thallusin in morphogenesis of the seaweed Ulva (Chlorophyta). J. Exp. Bot. 2020, 71, 3340–3349. [Google Scholar] [CrossRef]
- Favot, G.; Cunha, M.E.; Quental-Ferreira, H.; Ester, M.; Serrão, Á. Production of Ulva sp. in multitrophic aquaculture in earth ponds. Aquac. Fish Stud. 2019, 1, 1–8. [Google Scholar]
- Kamermans, P.; Malta, E.; Verschuure, J.M.; Lentz, L.F.; Schrijvers, L. Role of cold resistance and burial for winter survival and spring initiation of an Ulva spp. (Chlorophyta) bloom in a eutrophic lagoon (Veerse Meer lagoon, The Netherlands). Mar. Biol. 1998, 131, 45–51. [Google Scholar] [CrossRef]
- Sode, S.; Bruhn, A.; Balsby, T.J.S.; Larsen, M.M.; Gotfredsen, A.; Rasmussen, M.B. Bioremediation of reject water from anaerobically digested waste water sludge with macroalgae (Ulva lactuca, Chlorophyta). Bioresour. Technol. 2013, 146, 426–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsagkamilis, P.; Danielidis, D.; Dring, M.J.; Katsaros, C. Removal of phosphate by the green seaweed Ulva lactuca in a small-scale sewage treatment plant (Ios Island, Aegean Sea, Greece). J. Appl. Phycol. 2010, 22, 331–339. [Google Scholar] [CrossRef]
- Castelar, B.; Reis, R.P.; dos Santos Calheiros, A.C. Ulva lactuca and U. flexuosa (Chlorophyta, Ulvophyceae) cultivation in Brazilian tropical waters: Recruitment, growth, and ulvan yield. J. Appl. Phycol. 2014, 26, 1989–1999. [Google Scholar] [CrossRef]
- Castro, P.C.D.O. Assessing Key-Habitat Loss Due to Eutrophication in the Mondego and Mira Estuaries. Ph.D. Thesis, University of Coimbra, Coimbra, Portugal, 2006. [Google Scholar]
- Fonseca, J.C.; Pardal, M.A.; Azeiteiro, U.M.; Marques, J.C. Estimation of ecological exergy using weighing parameters determined from DNA contents of organisms—A case study. In Nutrients and Eutrophication in Estuaries and Coastal Waters; Springer: Dordrecht, The Netherlands, 2002; pp. 79–90. [Google Scholar]
- Rodrigues, F.A.A. Otimização das Condições de Cultivo de Ulva lactuca (Chlorophyta, Chlorophyceae) para Alimentação de Haliotis tuberculata. Master’s Thesis, University of Azores, Ponta Delgada, Portugal, 2016. [Google Scholar]
- Yaich, H.; Garna, H.; Besbes, S.; Paquot, M.; Blecker, C.; Attia, H. Effect of extraction conditions on the yield and purity of ulvan extracted from Ulva lactuca. Food Hydrocoll. 2013, 31, 375–382. [Google Scholar] [CrossRef]
- Chen, B.; McClements, D.J.; Decker, E.A. Design of foods with bioactive lipids for improved health. Annu. Rev. Food Sci. Technol. 2013, 4, 35–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Paul, V.J.; Luesch, H. Seaweed extracts and unsaturated fatty acid constituents from the green alga Ulva lactuca as activators of the cytoprotective Nrf2–ARE pathway. Free Radic. Biol. Med. 2013, 57, 141–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, Y.B. Ulva lactuca as bioindicator of metal contamination in intertidal waters in Hong Kong. Hydrobiologia 1990, 203, 73–81. [Google Scholar] [CrossRef]
- El-Sikaily, A.; El Nemr, A.; Khaled, A.; Abdelwehab, O. Removal of toxic chromium from wastewater using green alga Ulva lactuca and its activated carbon. J. Hazard. Mater. 2007, 148, 216–228. [Google Scholar] [CrossRef]
- Milinovic, J.; Rodrigues, C.; Diniz, M.; Noronha, J.P. Determination of total iodine content in edible seaweeds: Application of inductively coupled plasma-atomic emission spectroscopy. Algal Res. 2021, 53, 102149. [Google Scholar] [CrossRef]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [Green Version]
- Cotas, J.; Pacheco, D.; Araujo, G.S.; Valado, A.; Critchley, A.T.; Pereira, L. On the health benefits vs. risks of seaweeds and their constituents: The curious case of the polymer paradigm. Mar. Drugs 2021, 19, 164. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Fukumoto, M.; Nishida, H.; Ikeda, I.; Sugano, M. The interrelated effects of n-6/n-3 and polyunsaturated/saturated ratios of dietary fats on the regulation of lipid metabolism in rats. J. Nutr. 1989, 119, 1893–1899. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Kubo, K. Relationship between tissue lipid peroxidation and peroxidizability index after α-linolenic, eicosapentaenoic, or docosahexaenoic acid intake in rats. Br. J. Nutr. 2003, 89, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joint FAO/WHO. Expert consultation on fats and fatty acids in human nutrition. In Interim Summary of Conclusions and Dietary Recommendations on Total Fat & Fatty Acid; WHO: Geneva, Switzerland, 2008; Available online: https://www.who.int/nutrition/topics/FFA_summary_rec_conclusion.pdf (accessed on 3 February 2022).
- Rocha, C.P.; Pacheco, D.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. Seaweeds as valuable sources of essential fatty acids for human nutrition. Int. J. Environ. Res. Public Health 2021, 18, 4968. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID of Aquarium | Initial Biomass (g) | Closed Controlled Cultivation SGR | Aquarium SGR | Final SGR | Final Biomass (g) |
|---|---|---|---|---|---|
| A | 4.18 | 5.08% | 4.84% | 4.94% | 47.07 |
| B | 4.01 | 4.72% | 3.90% | 4.25% | 32.15 |
| C | 4.03 | 5.48% | 3.77% | 4.49% | 36.63 |
| Indoor total | 4.03 ± 0.08 | 5.09 ± 0.31 | 4.17 ± 0.48 | 4.56 ± 0.29 | 38.62 ± 6.25 |
| Tank cultivation | 878 | - | - | 4.37% | 2200 |
| FA | A | B | C | F | T | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C14:0 | 39.66 | ± | 3.23 | 35.79 | ± | 3.49 | 39.15 | ± | 2.71 | 48.34 | ± | 1.20 | 117.27 | ± | 3.11 |
| C16:0 | 1431.98 | ± | 107.33 | 1171.27 | ± | 35.27 | 949.87 | ± | 22.10 | 764.09 | ± | 44.19 | 922.94 | ± | 18.43 |
| C18:0 | 34.78 | ± | 8.89 | 34.45 | ± | 7.57 | 32.43 | ± | 6.54 | 31.27 | ± | 10.11 | 43.39 | ± | 2.73 |
| ∑ SFA | 1506.42 | ± | 99.78 | 1241.51 | ± | 42.74 | 1021.45 | ± | 26.23 | 843.70 | ± | 50.05 | 1083.59 | ± | 20.30 |
| C16:1 | 53.44 | ± | 3.24 | 35.87 | ± | 1.39 | 34.02 | ± | 0.46 | 35.64 | ± | 1.82 | 77.25 | ± | 1.54 |
| C18:1 | 496.17 | ± | 17.20 | 379.09 | ± | 3.51 | 344.95 | ± | 6.71 | 269.15 | ± | 16.34 | 441.69 | ± | 15.03 |
| ∑ MUFA | 549.60 | ± | 15.11 | 414.96 | ± | 3.31 | 378.986 | ± | 6.78 | 304.78 | ± | 18.13 | 518.94 | ± | 13.72 |
| C18:2 | 342.11 | ± | 20.05 | 322.11 | ± | 9.04 | 263.05 | ± | 7.01 | 145.67 | ± | 8.54 | 78.12 | ± | 1.48 |
| C16:3 | 52.69 | ± | 10.10 | 60.97 | ± | 6.10 | 46.59 | ± | 1.94 | 38.83 | ± | 1.49 | 16.77 | ± | 1.04 |
| C18:3 | 296.59 | ± | 5.06 | 359.80 | ± | 8.61 | 256.31 | ± | 8.56 | 437.42 | ± | 27.36 | 173.98 | ± | 7.60 |
| ∑ PUFA | 691.40 | ± | 12.67 | 742.87 | ± | 6.03 | 565.96 | ± | 14.97 | 621.92 | ± | 37.23 | 268.87 | ± | 9.95 |
| C16:4 | 59.50 | ± | 4.63 | 90.19 | ± | 7.29 | 59.78 | ± | 0.51 | 258.35 | ± | 10.46 | 121.36 | ± | 9.48 |
| C18:4 | 69.02 | ± | 4.31 | 79.99 | ± | 4.64 | 53.92 | ± | 1.82 | 319.02 | ± | 13.71 | 194.96 | ± | 11.82 |
| ∑ HUFA | 128.50 | ± | 8.69 | 170.18 | ± | 11.22 | 113.69 | ± | 2.10 | 577.37 | ± | 24.17 | 316.32 | ± | 21.28 |
| ∑ FA | 2875.92 | ± | 115.69 | 2569.51 | ± | 61.31 | 2080.08 | ± | 36.49 | 2347.77 | ± | 121.60 | 2187.72 | ± | 63.32 |
| Nutrients | A | B | C | F | T |
|---|---|---|---|---|---|
| PUFAs/SFA ratio | 0.91 | 1.07 | 1.06 | 1.78 | 1.02 |
| Total FAs | 0.03% | 0.03% | 0.02% | 0.03% | 0.02% |
| HUFA (EPA and DHA) (%) | 0.3% | 0.4% | 0.3% | 1.4% | 0.8% |
| Sugars (%) | 4.1% | 6.5% | 6.2% | 3.0% | 3.3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Poza, S.; Morais, T.; Leandro, A.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. A Comparative Study of the Fatty Acids and Monosaccharides of Wild and Cultivated Ulva sp. J. Mar. Sci. Eng. 2022, 10, 233. https://doi.org/10.3390/jmse10020233
García-Poza S, Morais T, Leandro A, Cotas J, Marques JC, Pereira L, Gonçalves AMM. A Comparative Study of the Fatty Acids and Monosaccharides of Wild and Cultivated Ulva sp. Journal of Marine Science and Engineering. 2022; 10(2):233. https://doi.org/10.3390/jmse10020233
Chicago/Turabian StyleGarcía-Poza, Sara, Tiago Morais, Adriana Leandro, João Cotas, João Carlos Marques, Leonel Pereira, and Ana M. M. Gonçalves. 2022. "A Comparative Study of the Fatty Acids and Monosaccharides of Wild and Cultivated Ulva sp." Journal of Marine Science and Engineering 10, no. 2: 233. https://doi.org/10.3390/jmse10020233