
Cytochrome b Sequencing as a Tool for Identification of Morphologically Similar Mediterranean Gobies Gobius incognitus and Gobius bucchichi (Actinopterygii: Gobiidae)


 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods

{kind=link}
{kind=link}
{kind=link}
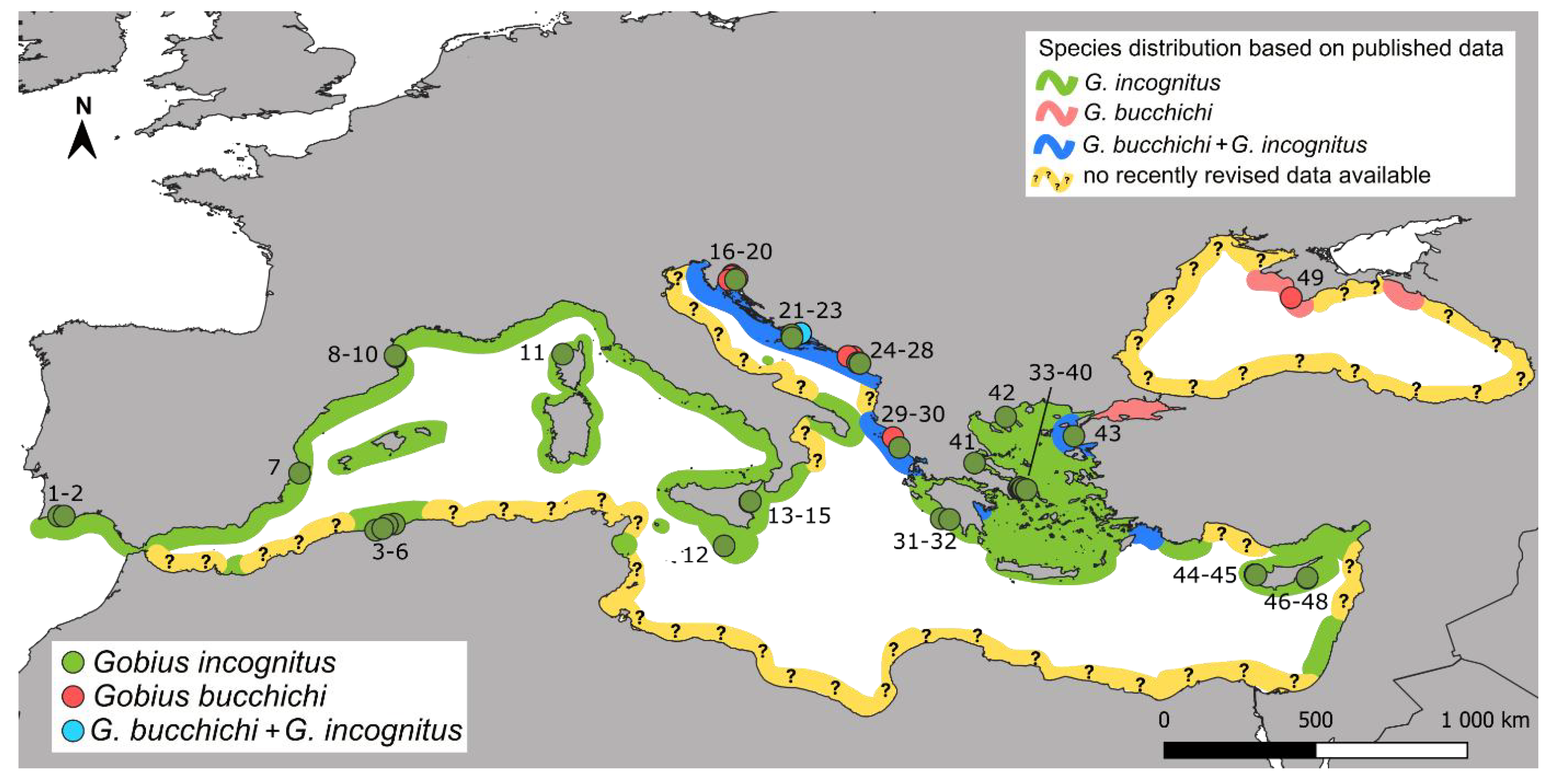
| Detected Species on Locality | Sea Subarea | Locality | Country | Coordinates | No. in Map | New Sequences | Published Sequences |
|---|---|---|---|---|---|---|---|
| Gobius incognitus | NE Atlantic | Gale | Portugal | 37.076972, −8.310722 | 1 | 7 | |
| Gobius incognitus | NE Atlantic | Sesmarias, Praia de Evaristo | Portugal | 37.074006, −8.303575 | 2 | 6 | |
| Gobius incognitus | African W Med | Bou Ismail | Algeria | 36.650767, 2.691153 | 3 | 4 | |
| Gobius incognitus | African W Med | Tipaza | Algeria | 36.594200, 2.449131 | 4 | 5 | |
| Gobius incognitus | African W Med | Algiers, Sidi Fredj | Algeria | 36.764291, 2.848401 | 5 | 17 | |
| Gobius incognitus | African W Med | Algiers, Ain Benian | Algeria | 36.802050, 2.898339 | 6 | 19 | |
| Gobius incognitus | Spanish W Med | Benidorm | Spain | 38.533982, −0.129395 | 7 | 4 | |
| Gobius incognitus | Gulf of Lion | Banyuls-sur-Mer | France | 42.481817, 3.136032 | 8 | 10 | 6 A |
| Gobius incognitus | Gulf of Lion | Banyuls-sur-Mer, Paulliles beach | France | 42.505759, 3.123662 | 9 | 7 | 3 A |
| Gobius incognitus | Gulf of Lion | Port Vendres | France | 42.523300, 3.109700 | 10 | 1 | |
| Gobius incognitus | Corsican shelf | Corsica, Lumio | France | 42.574444, 8.805278 | 11 | 25 | |
| Gobius incognitus | Maltese shelf | Gozo Island, Ramla | Malta | 36.061869, 14.284042 | 12 | 2 | |
| Gobius incognitus | Ionian Sea | Sicily, Aci Castello | Italy | 37.555419, 15.149383 | 13 | 9 | |
| Gobius incognitus | Ionian Sea | Sicily, Aci Trezza | Italy | 37.562758, 15.163661 | 14 | 9 | |
| Gobius incognitus | Ionian Sea | Sicily, Capo Mulini | Italy | 37.574014, 15.173408 | 15 | 8 | |
| Gobius bucchichi | Adriatic Sea | Kraljevica | Croatia | 45.267660, 14.562256 | 16 | 1 | 1 A |
| Gobius bucchichi | Adriatic Sea | Krk Island, Omišalj | Croatia | 45.218911, 14.550908 | 17 | 2 A | |
| Gobius bucchichi | Adriatic Sea | Krk Island, Glavotok | Croatia | 45.094134, 14.436694 | 18 | 1 B | |
| Gobius bucchichi | Adriatic Sea | Selce | Croatia | 45.152293, 14.718928 | 19 | 6 | 1 A |
| Gobius incognitus | Adriatic Sea | Krk Island, Sveti Marak | Croatia | 45.105944, 14.668500 | 20 | 2 | |
| Gobius bucchichi + Gobius incognitus | Adriatic Sea | Brač Island, Sumartin | Croatia | 43.285934, 16.878829 | 21 | 10 (5xG. inc., 5xG. buc.) | 6 A(G. inc.) |
| Gobius incognitus | Adriatic Sea | Hvar Island, Smočiguzica | Croatia | 43.233889, 16.574722 | 22 | 1 | |
| Gobius incognitus | Adriatic Sea | Hvar Island, Sveta Nedjelja | Croatia | 43.134433, 16.591381 | 23 | 14 | |
| Gobius bucchichi | Adriatic Sea | Boka Kotorska, Kostanjica | Montenegro | 42.485403, 18.669678 | 24 | 6 | |
| Gobius bucchichi | Adriatic Sea | Boka Kotorska, Strp | Montenegro | 42.503989, 18.669453 | 25 | 4 | |
| Gobius incognitus | Adriatic Sea | Budva, Jaz beach | Montenegro | 42.283836, 18.809034 | 26 | 1 | 2 A |
| Gobius incognitus | Adriatic Sea | Kamenovo | Montenegro | 42.273917, 18.887667 | 27 | 5 | |
| Gobius incognitus | Adriatic Sea | Sveti Stefan | Montenegro | 42.257180, 18.892118 | 28 | 5 A | |
| Gobius bucchichi | Ionian Sea | outflow of Butrint lagoon | Albania | 39.743614, 20.018654 | 29 | 1 | 5 A |
| Gobius incognitus | Ionian Sea | Sivota | Greece | 39.399800, 20.234664 | 30 | 2 | |
| Gobius incognitus | Ionian Sea | Romanos, Peloponnesos | Greece | 36.985361, 21.651634 | 31 | 1 | |
| Gobius incognitus | Ionian Sea | Petalidi, Peloponnesos | Greece | 36.959142, 21.934967 | 32 | 1 | |
| Gobius incognitus | Aegean Sea | Euboia Island, Petalii Islands | Greece | 38.014764, 24.280875 | 33 | 3 | |
| Gobius incognitus | Aegean Sea | Euboia Island, Marmari | Greece | 38.048808, 24.318328 | 34 | 4 | |
| Gobius incognitus | Aegean Sea | Euboia Island, Kalamitsi beach | Greece | 37.971372, 24.365764 | 35 | 2 | |
| Gobius incognitus | Aegean Sea | Euboia Island, Mnima penninsula | Greece | 37.978775, 24.399581 | 36 | 4 | |
| Gobius incognitus | Aegean Sea | Euboia Island, Agia Pelagia Islet | Greece | 37.997008, 24.398186 | 37 | 1 | |
| Gobius incognitus | Aegean Sea | Euboia Island, Karystos | Greece | 38.009667, 24.430804 | 38 | 3 | |
| Gobius incognitus | Aegean Sea | Euboia Island, Kastri Beach | Greece | 37.973022, 24.537658 | 39 | 1 | |
| Gobius incognitus | Aegean Sea | Euboia Island, Mirmigia rocks | Greece | 37.972336, 24.548042 | 40 | 1 | |
| Gobius incognitus | Aegean Sea | Raches | Greece | 38.876144, 22.785700 | 41 | 3 | |
| Gobius incognitus | Aegean Sea | Gavradia | Greece | 40.440278, 23.839722 | 42 | 18 | |
| Gobius incognitus | Aegean Sea | between Geyikli and Dalyan * | Turkey | 39.789722, 26.156111 | 43 | 22 | |
| Gobius incognitus | Levantine Sea | Akamas, Fontana | Cyprus | 35.090108, 32.303844 | 44 | 3 | |
| Gobius incognitus | Levantine Sea | Akamas, George Island | Cyprus | 35.074958, 32.335222 | 45 | 8 | |
| Gobius incognitus | Levantine Sea | Cavo Greco, Tunnel—cave | Cyprus | 34.963592, 34.073078 | 46 | 7 | |
| Gobius incognitus | Levantine Sea | Cavo Greco, chappel | Cyprus | 34.976053, 34.076781 | 47 | 2 | |
| Gobius incognitus | Levantine Sea | Cavo Greco, Cyclops cave | Cyprus | 34.985631, 34.076928 | 48 | 1 | |
| Gobius bucchichi | Black Sea | Sevastopol | Ukraine | 44.494094, 33.533650 | 49 | 1 |
3. Results
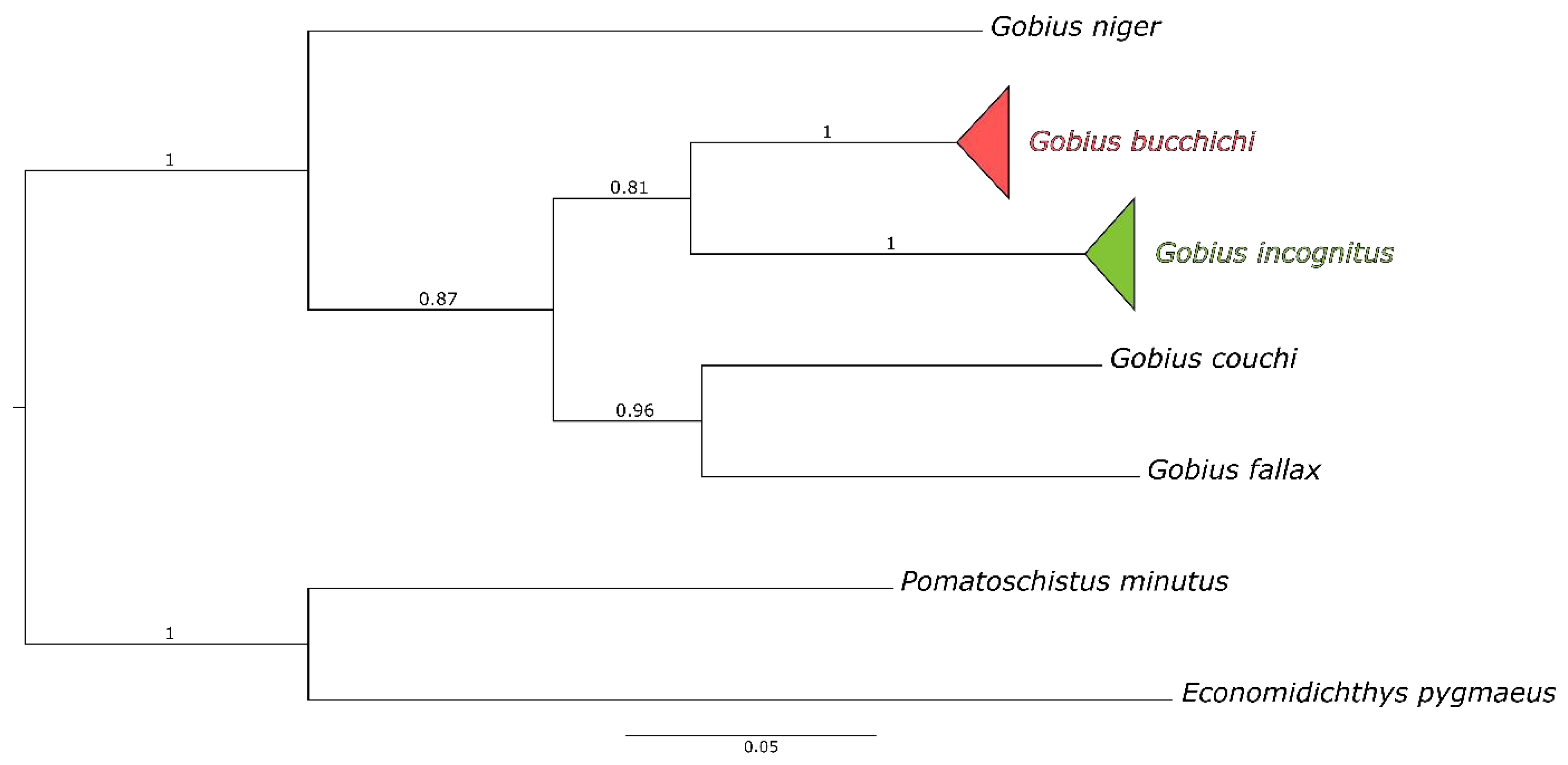
3.1. Genetic Distinctiveness
3.2. Distribution
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parenti, P. A checklist of the gobioid fishes of the world (Percomorpha: Gobiiformes). Iran. J. Ichthyol. 2021, 8, 1–480. [Google Scholar] [CrossRef]
- Kovačić, M. Checklist of gobies (Teleostei: Gobiidae) of the Mediterranean Sea and a key for species identification. Zootaxa 2020, 4877, 75–101. [Google Scholar] [CrossRef]
- McCraney, W.T.; Thacker, C.E.; Alfaro, M.E. Supermatrix phylogeny resolves goby lineages and reveals unstable root of Gobiaria. Mol. Phylogenet. Evol. 2020, 151, 106862. [Google Scholar] [CrossRef] [PubMed]
- Iglésias, S.P.; Vukić, J.; Sellos, D.Y.; Soukupová, T.; Šanda, R. Gobius xoriguer, a new offshore Mediterranean goby (Gobiidae), and phylogenetic relationships within the genus Gobius. Ichthyol. Res. 2021, 68, 445–459. [Google Scholar] [CrossRef]
- Kovačić, M.; Ordines, F.; Schliewen, U.K. A new species of Speleogobius (Teleostei: Gobiidae) from the Western Mediterraenean Sea. Zootaxa 2016, 4066, 301–310. [Google Scholar] [CrossRef]
- Kovačić, M.; Ordines, F.; Schliewen, U.K. A new species of Buenia (Teleostei: Gobiidae) from the western Mediterranean Sea, with the description of this genus. Zootaxa 2017, 4250, 447–460. [Google Scholar] [CrossRef]
- Kovačić, M.; Ordines, F.; Schliewen, U.K. A new species of Buenia (Perciformes: Gobiidae) from the western Mediterranean slope bottoms, the redescription of Buenia jeffreysi and the first Balearic record of Buenia affinis. Zootaxa 2018, 4392, 267–288. [Google Scholar] [CrossRef]
- Kovačić, M.; Ordines, F.; Ramirez-Amaro, S.; Schliewen, U.K. Gymnesigobius medits (Teleostei: Gobiidae), a new gobiid genus and species from the western Mediterranean slope bottoms. Zootaxa 2019, 4651, 513–530. [Google Scholar] [CrossRef]
- Kovačić, M.; Šanda, R.; Čekovská, K.; Soukupová, T.; Vukić, J. Zebrus pallaoroi sp. nov.: A new species of goby (Actinopterygii: Gobiidae) from the Mediterranean Sea with a dna-based phylogenetic analysis of the Gobius-lineage. Contrib. Zool. 2021, 90, 285–317. [Google Scholar] [CrossRef]
- Kovačić, M.; Glavičić, I. The first Adriatic finding of Speleogobius llorisi (Actinopterygii: Gobiiformes: Gobiidae). Acta Ichthyol. Piscat. 2019, 49, 181–184. [Google Scholar] [CrossRef]
- Dulčić, J.; Lepen Pleić, I.; Zorica, B.; Bušelić, I.; Šestanović, M.; Kovačić, M. Fish larvae DNA barcoding indicated the potential appearance of rare species: Buenia massutii Kovačić, Ordines, and Schliewen, 2017 in the Adriatic Sea. Acta Adriat. 2022, 63, 45–52. [Google Scholar] [CrossRef]
- Seyhan Öztürk, D.; Oruç, A.Ç. The new distribution records of recently described gobies: Aegean Sea records for Pomatoschistus nanus Engin & Seyhan OztUrk, 2017 and easternmost records of Buenia massutii Kovačić, Ordines & Schliewen, 2017. J. Mar. Biol. Assoc. U. K. 2023, 103, e40. [Google Scholar] [CrossRef]
- Kovačić, M.; Froglia, C. The Adriatic record of Gobius xoriguer Iglésias, Vukić & Šanda, 2021 and additional records of Vanneaugobius dollfusi Brownell, 1978 (Actinopterygii: Gobiiformes: Gobiidae). Cybium 2023, 47, 193–196. [Google Scholar] [CrossRef]
- Kovačić, M.; Ramírez-Amaro, S.; Farriols, M.T.; Ordines, F. The second record of Gymnesigobius medits Kovačić, Ordines, Ramirez-Amaro & Schliewen, 2019, the deepest benthic gobiiform species, and the additional records of Gobius xoriguer Iglésias, Vukić & Šanda, 2021 (Actinopterygii: Gobiiformes: Gobiidae). Fishes 2023, 8, 331. [Google Scholar] [CrossRef]
- Kovačić, M.; Šanda, R.; Vukić, J.; Renoult, J.P.; Falzon, M.-A. New records of the recently described Pomatoschistus nanus Engin & Seyhan, 2017 (Teleostei: Gobiidae). Cybium 2023, 2017, 1–4. [Google Scholar] [CrossRef]
- Kovačić, M.; Šanda, R.; Kirinčić, M.; Zanella, D. Geographic distribution of gobies (Gobiidae) in the Adriatic Sea with thirteen new records for its southern part. Cybium 2012, 36, 435–445. [Google Scholar] [CrossRef]
- Miller, P.J. Fishes of the North-eastern Atlantic and the Mediterranean. In Gobiidae; Whitehead, P.J.P., Bauchot, M.-L., Hureau, J.-C., Nielsen, J., Tortonese, E., Eds.; Unesco: Paris, France, 1986; pp. 1019–1085. [Google Scholar]
- Kovačić, M.; Šanda, R. A new species of Gobius (Perciformes: Gobiidae) from the Mediterranean Sea and the redescription of Gobius bucchichi. J. Fish Biol. 2016, 88, 1104–1124. [Google Scholar] [CrossRef]
- Kovačić, M.; Renoult, J.P.; Pillon, R.; Svensen, R.; Bogorodsky, S.V.; Engin, S.; Louisy, P. Identification of Mediterranean marine gobies (Actinopterygii: Gobiidae) of the continental shelf from photographs of in situ individuals. Zootaxa 2022, 5144, 1–103. [Google Scholar] [CrossRef]
- Kovačić, M.; Renoult, J.P.; Pillon, R.; Bilecenoglu, M.; Tiralongo, F.; Bogorodsky, S.V.; Engin, S.; Kovtun, O.; Louisy, P.; Patzner, R.A.; et al. The delimitation of geographic distributions of Gobius bucchichi and Gobius incognitus (Teleostei: Gobiidae). J. Mar. Sci. Eng. 2023, 11, 516. [Google Scholar] [CrossRef]
- Glavičić, I.; Kovačić, M.; Soldo, A.; Schliewen, U. A quantitative assessment of the diel influence on the cryptobenthic fish assemblage of the shallow Mediterranean infralittoral zone. Sci. Mar. 2020, 84, 49–57. [Google Scholar] [CrossRef]
- Tiralongo, F.; Pillon, R. New distributional records of Gobius bucchichi (Pisces, Gobiidae) from the Mediterranean Sea and in situ comparisons with Gobius incognitus. Ann. za Istrske Mediteranske Stud. Ser. Hist. Sociol. 2020, 30, 215–220. [Google Scholar] [CrossRef]
- Tiralongo, F.; Messina, G.; Lombardo, B.M. First data on habitat preference, diet and length-weight relationship of Gobius incognitus Kovačić & Šanda, 2016 (Pisces: Gobiidae). Acta Adriat. 2020, 61, 67–78. [Google Scholar] [CrossRef]
- Soldo, A.; Glavičić, I.; Kovačić, M. Combining methods to better estimate total fish richness on temperate reefs: The case of a Mediterranean coralligenous cliff. J. Mar. Sci. Eng. 2021, 9, 670. [Google Scholar] [CrossRef]
- Bilecenoğlu, M.; Yokeş, M.B. Distribution of two closely allied gobies, Gobius bucchichi Steindachner, 1870 and Gobius incognitus Kovačić & Šanda, 2016, along the Turkish coast. FishTaxa 2022, 23, 47–53. [Google Scholar]
- Kovačić, M.; Kovtun, O.A. The first record of Gobius incognitus (Actinopterygii: Gobiiformes: Gobiidae) from Malta. Acta Adriat. 2022, 63, 53–58. [Google Scholar] [CrossRef]
- Renoult, J.P.; Pillon, R.; Kovačić, M.; Louisy, P. Frontiers in Fishwatching Series-Gobies of the North-eastern Atlantic and the Mediterranean: Gobius and Thorogobius. Les Cah. Fond. Biotope 2022, 37, 1–237. [Google Scholar]
- Spatafora, D.; Cattano, C.; Aglieri, G.; Quattrocchi, F.; Turco, G.; Quartararo, G.; Dudemaine, J.; Calosi, P.; Milazzo, M. Limited behavioural effects of ocean acidification on a Mediterranean anemone goby (Gobius incognitus) chronically exposed to elevated CO2 levels. Mar. Environ. Res. 2022, 181, 105758. [Google Scholar] [CrossRef]
- Francour, P.; Bilecenoglu, M.; Bariche, M.; Tunesi, L.; Goren, M. Gobius bucchichi. The IUCN Red List of Threatened Species 2011. Available online: https://www.iucnredlist.org/species/194873/8911208 (accessed on 20 October 2023). [CrossRef]
- Milana, V.; Šanda, R.; Vukić, J.; Ciccotti, E.; Riccato, F.; Petrosino, G.; Rossi, A.R. Far from home: Genetic variability of Knipowitschia sp. from Italy revealed unexpected species in coastal lagoons of the Tyrrhenian coast. Estuar. Coast. Shelf Sci. 2021, 251, 107260. [Google Scholar] [CrossRef]
- Vukić, J.; Kovačić, M.; Zogaris, S.; Šanda, R. Rediscovery of Knipowitschia goerneri and its molecular relationships with other European northern Mediterranean Knipowitschia species (Teleostei: Gobiidae). Ichthyol. Explor. Freshwaters 2016, 26, 363–372. [Google Scholar]
- Vukić, J.; Ulqini, D.; Šanda, R. Occurrence of Knipowitschia goerneri Ahnelt, 1991 (Gobiidae) in southern Albania confirmed with molecular tools. J. Appl. Ichthyol. 2017, 33, 284–290. [Google Scholar] [CrossRef]
- Agorreta, A.; San Mauro, D.; Schliewen, U.; Van Tassell, J.L.; Kovačić, M.; Zardoya, R.; Rüber, L. Molecular phylogenetics of Gobioidei and phylogenetic placement of European gobies. Mol. Phylogenet. Evol. 2013, 69, 619–633. [Google Scholar] [CrossRef]
- Neilson, M.E.; Stepien, C.A. Escape from the Ponto-Caspian: Evolution and biogeography of an endemic goby species flock (Benthophilinae: Gobiidae: Teleostei). Mol. Phylogenet. Evol. 2009, 52, 84–102. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, D.A.; Sorokin, P.A.; Vasil’ev, V.P.; Chernova, N.V.; Vasil’eva, E.D. Reconstruction of phylogenetic relations of Ponto-Caspian gobies (Gobiidae, Perciformes) based on mitochondrial genome variation and some problems of their taxonomy. J. Ichthyol. 2013, 53, 702–712. [Google Scholar] [CrossRef]
- Tougard, C.; Vukić, J.; Ahnelt, H.; Buj, I.; Kovačić, M.; Moro, G.A.; Tutman, P.; Šanda, R. Quaternary climatic cycles promoted (re)colonization and diversification events in Adriatic sand gobies. J. Zool. Syst. Evol. Res. 2021, 59, 1037–1052. [Google Scholar] [CrossRef]
- Čekovská, K.; Šanda, R.; Eliášová, K.; Kovačić, M.; Zogaris, S.; Pappalardo, A.M.; Soukupová, T.; Vukić, J. Population genetic diversity of two marine gobies (Gobiiformes: Gobiidae) from the North-eastern Atlantic and the Mediterranean Sea. J. Mar. Sci. Eng. 2020, 8, 792. [Google Scholar] [CrossRef]
- Gysels, E.S.; Hellemans, B.; Pampoulie, C.; Volckaert, F.A.M. Phylogeography of the common goby, Pomatoschistus microps, with particular emphasis on the colonization of the Mediterranean and the North Sea. Mol. Ecol. 2004, 13, 403–417. [Google Scholar] [CrossRef]
- Larmuseau, M.H.D.; Huyse, T.; Vancampenhout, K.; Van Houdt, J.K.J.; Volckaert, F.A.M. High molecular diversity in the rhodopsin gene in closely related goby fishes: A role for visual pigments in adaptive speciation? Mol. Phylogenet. Evol. 2010, 55, 689–698. [Google Scholar] [CrossRef]
- Boissin, E.; Hoareau, T.B.; Berrebi, P. Effects of current and historic habitat fragmentation on the genetic structure of the sand goby Pomatoschistus minutus (Osteichthys, Gobiidae). Biol. J. Linn. Soc. 2011, 102, 175–198. [Google Scholar] [CrossRef]
- Mejri, R.; Arculeo, M.; Ben Hassine, O.K.; Lo Brutto, S. Genetic architecture of the marbled goby Pomatoschistus marmoratus (Perciformes, Gobiidae) in the Mediterranean Sea. Mol. Phylogenet. Evol. 2011, 58, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Tougard, C.; Folly, J.; Berrebi, P. New light on the evolutionary history of the common goby (Pomatoschistus microps) with an emphasis on colonization processes in the Mediterranean Sea. PLoS ONE 2014, 9, e91576. [Google Scholar] [CrossRef] [PubMed]
- Seyhan-Ozturk, D.; Engin, S. Genetic diversity of marbled goby populations in the Anatolian coasts of the north-eastern Mediterranean. J. Mar. Biol. Assoc. U. K. 2021, 101, 419–429. [Google Scholar] [CrossRef]
- Šanda, R.; Vukić, J.; Choleva, L.; Křížek, J.; Šedivá, A.; Shumka, S.; Wilson, I.F. Distribution of loach fishes (Cobitidae, Nemacheilidae) in Albania, with genetic analysis of populations of Cobitis ohridana. Folia Zool. 2008, 57, 42–50. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Villesen, P. FaBox: An online toolbox for FASTA sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Šanda, R.; Kovačić, M. First record of Gobius couchi (Gobiidae) in the Ionian Sea. Cybium 2009, 33, 249–250. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree Version 1.4.4. 2018. Available online: https://github.com/rambaut/figtree/releases (accessed on 2 February 2019).
- Inkscape Project. Inkscape Version 0.92.3. Available online: https://inkscape.org (accessed on 10 June 2022).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Francisco, S.M.; Castilho, R.; Lima, C.S.; Almada, F.; Rodrigues, F.; Šanda, R.; Vukić, J.; Pappalardo, A.M.; Ferrito, V.; Robalo, J.I. Genetic hypervariability of a Northeastern Atlantic venomous rockfish. PeerJ 2021, 9, e11730. [Google Scholar] [CrossRef]
- Reichenbacher, B.; Gregorová, R.; Holcová, K.; Šanda, R.; Vukić, J.; Přikryl, T. Discovery of the oldest Gobius (Teleostei, Gobiiformes) from a marine ecosystem of Early Miocene age. J. Syst. Palaeontol. 2018, 16, 493–513. [Google Scholar] [CrossRef]
- Webb, C.J. Systematics of the Pomatoschistus minutus complex (Teleostei: Gobioidei). Philos. Trans. R. Soc. Lond. B Biol. Sci. 1980, 291, 201–241. [Google Scholar] [CrossRef]
- Berrebi, P.; Trébuchon, M. Distribution and hybridization of two sedentary gobies (Pomatoschistus microps and Pomatoschistus marmoratus) in the lagoons of southern France. Sci. Mar. 2020, 84, 355–367. [Google Scholar] [CrossRef]
- Horvatić, S.; Malavasi, S.; Vukić, J.; Šanda, R.; Marčić, Z.; Ćaleta, M.; Lorenzoni, M.; Mustafić, P.; Buj, I.; Onorato, L.; et al. Correlation between acoustic divergence and phylogenetic distance in soniferous European gobiids (Gobiidae; Gobius lineage). PLoS ONE 2021, 16, e0260810. [Google Scholar] [CrossRef] [PubMed]
- Horvatić, S.; Parmentier, E.; Malavasi, S.; Clara Amorim, M.P.; Fonseca, P.J.; Zanella, D. Endemic fish calling: Acoustics and reproductive behaviour of the Neretva dwarf goby Orsinigobius croaticus. Ecol. Evol. 2023, 13, e10673. [Google Scholar] [CrossRef] [PubMed]
- Bariche, M.; Fricke, R. The marine ichthyofauna of Lebanon: An annotated checklist, history, biogeography, and conservation status. Zootaxa 2020, 4775, 1–157. [Google Scholar] [CrossRef] [PubMed]
- Soldo, A.; Bakiu, R. Checklist of marine fishes of Albania. Acta Adriat. 2021, 62, 63–73. [Google Scholar] [CrossRef]
- Bitar, G.; Badreddine, A. An updated checklist of the marine fishes in Lebanon. An answer to Bariche and Fricke (2020): “The marine ichthyofauna of Lebanon: An annotated checklist, history, biogeography, and conservation status”. Zootaxa 2021, 5010, 1–128. [Google Scholar] [CrossRef]
- Elbaraasi, H.; Elabar, B.; Elaabidi, S.; Bashir, A.; Elsilini, O.; Shakman, E.; Azzurro, E. Updated checklist of bony fishes along the Libyan coast (southern Mediterranean Sea). Mediterr. Mar. Sci. 2019, 20, 90–105. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Soliman, I.A. Checklist of the bony fish species in the Mediterranean waters of Egypt. Bull. Natl. Inst. Oceanogr. Fish. 1996, 22, 43–57. [Google Scholar]
- Bouchereau, J.L.; Durel, J.S.; Guelorget, O.; Louali, L.R. L’ichtyofaune dans l’organisation biologique d’un système paralique marocain: La lagune de Nador (in french). Mar. Life 2000, 10, 69–76. [Google Scholar]
- Ali, M.F. An updated Checklist of Marine fishes from Syria with an emphasis on alien species. Mediterr. Mar. Sci. 2018, 19, 388–393. [Google Scholar] [CrossRef]
- Keskin, Ç. A review of fish fauna in the Turkish Black Sea. Karadeniz balık faunası. J. Black Sea/Mediterr. Environ. 2010, 16, 195–210. [Google Scholar]
- Vasiľeva, E.D. Fishes of the Black Sea. Key to Marine, Brackish–Water, Euryhaline and Anadromous Species with Color Illustrations, Collected by S.V. Bogorodsky; VNIRO Publishing: Moscow, Russia, 2007; pp. 1–237. (In Russian) [Google Scholar]
- Parin, N.V.; Evseenko, S.A.; Vasiľeva, E.D. Fishes of Russian Seas: Annotated Catalogue; KMK Scientific Press Ltd.: Moscow, Russia, 2014; pp. 1–733. [Google Scholar]
- Manilo, L.G. Ryby Semeystva Bychkovye (Perciformes, Gobiidae) Morskikh i Solonovatykh vod Ukrainy; Naukova Dumka: Kyiv, Ukraine, 2014; pp. 1–243. (In Russian) [Google Scholar]
- Boltachev, A.R.; Karpova, E.P. Marine Fishes of the Crimean Peninsula, 2nd ed.; Biznes-Inform: Simferopol, Ukraine, 2017; pp. 1–376. (In Russian) [Google Scholar]
- Vassilev, M.; Apostolou, A.; Velkov, B.; Dobrev, D.; Zarev, V. Atlas of the Gobies (Gobiidae) in Bulgaria; Institute of Biodiversity and Ecosystem Research, Bulgarian Academy of Sciences: Sofia, Bulgaria, 2012; pp. 1–114. [Google Scholar]
- Giantsis, I.A.; Tokamani, M.; Triantaphyllidis, G.; Tzatzani, S.; Chatzinikolaou, E.; Toros, A.; Bouchorikou, A.; Chatzoglou, E.; Miliou, H.; Sarantopoulou, J.; et al. Development of multiplex PCR and melt–curve analysis for the molecular identification of four species of the Mullidae family, available in the market. Genes 2023, 14, 960. [Google Scholar] [CrossRef]



| G. incognitus | G. bucchichi | G. couchi | G. falax | G. niger | |
|---|---|---|---|---|---|
| G. incognitus (258) | 0.63 (±0.086) | 0.8 | 0.9 | 1.1 | 1.1 |
| G. bucchichi (30) | 13 | 0.68 (±0.11) | 1 | 1.1 | 1.2 |
| G. couchi (1) | 16.7 | 14.9 | N/A | 1.1 | 1.1 |
| G. falax (1) | 16.6 | 15.3 | 14.3 | N/A | 1.2 |
| G. niger (1) | 19.4 | 18 | 18.9 | 19.7 | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čekovská, K.; Šanda, R.; Ašenbrenerová, E.; Kassar, A.; Zogaris, D.; Pappalardo, A.M.; Tarkan, A.S.; Vasil’eva, E.; Santos, D.; Vukić, J. Cytochrome b Sequencing as a Tool for Identification of Morphologically Similar Mediterranean Gobies Gobius incognitus and Gobius bucchichi (Actinopterygii: Gobiidae). J. Mar. Sci. Eng. 2023, 11, 2289. https://doi.org/10.3390/jmse11122289
Čekovská K, Šanda R, Ašenbrenerová E, Kassar A, Zogaris D, Pappalardo AM, Tarkan AS, Vasil’eva E, Santos D, Vukić J. Cytochrome b Sequencing as a Tool for Identification of Morphologically Similar Mediterranean Gobies Gobius incognitus and Gobius bucchichi (Actinopterygii: Gobiidae). Journal of Marine Science and Engineering. 2023; 11(12):2289. https://doi.org/10.3390/jmse11122289
Chicago/Turabian StyleČekovská, Katarína, Radek Šanda, Eva Ašenbrenerová, Abderrahmane Kassar, Dimitris Zogaris, Anna Maria Pappalardo, Ali Serhan Tarkan, Ekaterina Vasil’eva, David Santos, and Jasna Vukić. 2023. "Cytochrome b Sequencing as a Tool for Identification of Morphologically Similar Mediterranean Gobies Gobius incognitus and Gobius bucchichi (Actinopterygii: Gobiidae)" Journal of Marine Science and Engineering 11, no. 12: 2289. https://doi.org/10.3390/jmse11122289