
Chara zeylanica J.G.Klein ex Willd. (Charophyceae, Charales, Characeae): First European Record from the Island of Sardinia, Italy


1
Independent Researcher, 26121 Oldenburg, Germany
2
Aquatic Ecology, Institute of Biosciences, University of Rostock, 18059 Rostock, Germany
*
Author to whom correspondence should be addressed.
Plants 2021, 10(10), 2069; https://doi.org/10.3390/plants10102069
Submission received: 27 July 2021
/
Revised: 23 September 2021
/
Accepted: 24 September 2021
/
Published: 30 September 2021
(This article belongs to the Special Issue Macrophytes in Inland Waters: From Knowledge to Management)
Abstract
:The first record of a species belonging to the genus Chara L. subgenus Chara R.D.Wood section Grovesia R.D.Wood subsect. Willdenowia R.D.Wood from Europe is presented here, thus challenging the interpretation of its distribution pattern as an intertropical group of charophytes. The morphological characters of the specimens, as well as the results of a phylogenetic analysis, clearly identified them as Chara zeylanica J.G.Klein ex Willd. Although the subsection Willdenowia has yet to receive a thorough taxonomic treatment, a discussion of its relationship to other taxa of this subsection is provided despite the lack of a commonly agreed upon taxonomic concept. The ecological conditions of the Sardinian site of C. zeylanica are presented. Moreover, the status of and threats to this taxon, and hypotheses regarding potential pathways through which it reached Europe, are discussed.
1. Introduction
Charophytes are morphologically complex macrophytic green algae with a worldwide distribution. Because they are close relatives of the earliest land plants [1], they have attracted growing scientific interest in recent decades. However, in addition to becoming a subject of academic interest, charophytes play a major role in bioindication systems due to their species-specific pattern of niche occupation [2,3]. Moreover, Characeae are among the most threatened groups of organisms on earth [4,5,6], and have thus been targeted by nature conservation actions [7,8,9]. As charophytes occur in an astonishingly wide variety of habitats, ranging from ultraoligotrophic freshwater to hypersaline and hypertrophic environments, their presence is often measured in water quality assessments and other related fields [10,11].
For the development of such bioindication systems, having comprehensive and reliable knowledge about the habitat preferences and distribution patterns of the individual species is essential, as is the accurate identification of charophyte species, and the formulation of a sound taxonomic concept. In recent decades, a large number of studies have attempted to fulfil these requirements [12,13,14,15,16,17,18,19]. As a result, our knowledge about the biogeography of charophytes has increased substantially. However, whereas in the past site-specific information about the occurrence of the individual species was provided [20], recent treatments have led to the development of large-scale distribution grid maps and detailed descriptions of the species’ preferred habitat conditions [21].
For several species, a strong correspondence between the distribution range and the niche structure was found. For example, the strictly circumpolar distribution of Tolypella normaniana Nordst. can be explained by its temperature preference (cold-stenothermic). Moreover, it has been shown that species such as Chara vulgaris L. or C. braunii C.C.Gmelin occur in a broad range of habitats on all continents, except for Antarctica [22].
However, unresolved questions regarding charophytes have hampered the development of general bioindication schemes that are also applicable outside of the reference regions, which have mainly been restricted to specific geographic scales [3]. One of these questions related to the absence of subsection Willdenowia R.D.Wood (section Grovesia R.D.Wood) in Europe is dealt with here, as we provide the first record of the presence of Chara zeylanica J.G.Klein ex Willd. in Europe from the Mediterranean island of Sardinia (Italy).
In a global taxonomic treatment of charophytes by Wood [23,24], the genus Chara L. was divided into five sections with a total of eight subsections. Subsection Willdenowia comprises diplostephanous species with triplostichous cortication and a completely ecorticated basal branchlet segment. According to Wood [23,24]), this subsection includes just one species, Chara zeylanica, which has several varieties and forms. This approach was not universally accepted because it combined A) monoecious and dioecious taxa, B) monoecious taxa with sejoined and conjoined gametangia, and C) taxa with tetra- and octoscutate antheridia [23]. However, several authors used Wood’s concept as a basis for investigating the distribution pattern of subsect. Willdenowia, and came to the conclusion that it can best be described as an intertropical taxon [25,26]. On the other hand, as distinct patterns of the distribution of subspecies and varieties of Chara zeylanica sensu Wood [23] emerged, a fine-resolution taxonomic treatment of subsection Willdenowia was clearly needed for biogeographical purposes [25,27]. In an approach designed to overcome the problems caused by Wood’s taxonomic concept, van Raam [28,29] presented an alternative view in which subsect. Willdenowia was divided into 20 species that were mainly distinguished by the abovementioned criteria of gametangia position, antheridia morphology, and sexuality.
However, irrespective of which concept was applied, neither Chara zeylanica nor any other taxon of subsect. Willdenowia has previously been recorded anywhere in Europe, even though numerous investigations of charophytes have been performed throughout the Mediterranean area in recent decades [30,31,32,33,34,35,36]. Chara zeylanica occurs mainly in tropical and subtropical regions of the world [23,37,38,39,40,41,42,43]. As it is an “intertropical taxon”, the first record of the presence of C. zeylanica in Europe could be considered a surprise. On the other hand, Corillion and Guerlesquin [26] and Proctor et al. [25] have reported, taxa of subsect. Willdenowia have been found in North America at up to 45° N under climatic conditions comparable to those in Northern Europe. There are historical records of the presence of the species from Egypt and Israel [26,44], as well as reports of extinct occurrences from Algeria [45]. Consequently, limitations other than climatic conditions should be responsible for the failure to observe the presence of taxa of subsect. Willdenowia in Europe, which is a well-investigated region that certainly cannot be regarded as undersampled. Recent records of the presence of non-native charophyte species with predominantly tropical and subtropical distributions—such as reports of the presence of Chara fibrosa C.Agardh ex Bruzelius ssp. benthamii (A.Braun) Zaneveld or Chara c.f. chrysospora J.Groves and Stevens in rice fields, lakes, and an artificial stormwater retention pond in Southern France, Italy, and Crete, respectively [32,46,47,48]—indicate that the climatic conditions in the Mediterranean area are suitable for the establishment of intertropical taxa.
The main aim of this study is to document the first record of the presence of Chara zeylanica in Europe, and the morphological features of the Sardinian specimens we collected. Moreover, this study contributes to knowledge about the taxonomic classification, the ecological requirements, and the geographic distribution of this mainly tropical and subtropical species. To support our morphological analysis, we used rbcL and matK barcodes, as previous barcoding studies have shown that a combination of these sequences is suitable for investigating species of the genus Chara [49,50,51,52].
2. Results
2.1. Ecology
Chara zeylanica can colonize a broad range of both brackish and freshwater habitats throughout the tropical and subtropical zones of the world. These habitats include permanent and temporary bodies of water, such as lakes, ponds, pools, ditches, temporarily flooded wetlands, canals, rice fields, and retention ponds [26,38,40,41,42,43]. Few of the existing hydrochemical datasets cover a spectrum ranging from low-impacted waterbodies with total P-concentrations below 20 µg L−1 and total N-concentrations between 0.425 and 1.9 mg L−1 [43] to eutrophic habitats [37]. According to Muller et al. [45], C. zeylanica needs temperatures of approximately 25 °C for fructification.
The only European site (reported here for the first time) where the presence of C. zeylanica has been detected is at Cala Fuili, which is located north of Orosei on the east coast of Sardinia, Italy (coordinates: 40°25′03″ N, 9°46′13″ E; coordinate system WGS 84) (Figure 1). The specimens were found in September 2019 at a depth of about 1 m, mainly in sandy to silty places with stony substrate, in a shallow and probably permanent small stream located close to the beach, or 110 m from the Mediterranean Sea. The specific site where the C. zeylanica specimens were found was situated directly next to a bridge (Figure 1, left image below), and was therefore disturbed by the structure. By contrast, the neighbouring stream sections and landscape areas can be considered semi-natural habitats. The small population of C. zeylanica was observed to have high fertility, with ripe antheridia, oogonia, and oospores. The nutrient conditions at the sampling date were as follows: NH4-N 0.108 mg L−1, NO3-N 0.279 mg L−1, total N 1.143 mg L−1, PO4-P 0.073 mg L- 1, and total P 0.137 mg L−1. The water hardness was 26.4 °dH (Ca 62.2 mg L−1, Mg 76.8 mg L- 1), pH 8.3. Although the salinity at the sampling date was 1.9, the salinity of the site probably varies because it is close to the coast. At the same site in May 2016, a salinity level of 4.4 was recorded and the Cl concentration was found to be 2819 mg L−1, instead of 1290 mg L- 1, as measured in September 2019.
2.2. Morphological Description
The specimens are 30–60 cm long, erect and straight, stout, fresh to greyish green, and slightly incrusted (Figure 2). The main axis diameter is 589–1076 µm with a mean value of 844 µm, slightly (0–4) branched. Most of the internodes are 4.4–8.0 cm long, and are usually much longer (up to 4 ×) than the branchlets. The uppermost 1–2 internodes are only 0.5–2.2 cm long, and are generally shorter than the adjacent branchlets. The cortex is usually triplostichous, and is rarely (partly) diplostichous and tylacanthous to isostichous (Figure 2D). Single, acute, thin, and needle-like spines are observed on the young internodes, and rarely on the older internodes. These spines can vary in length (182–1468 µm long) even on the same plant, and mainly point downwards (Figure 2D). The stipulodes are acute, elongated, and well developed. They are arranged in two regular tiers with two pairs per branchlet (Figure 2E). The upper stipulodes are longer than the lower ones. As the upper stipulodes are 515–1045 µm long (a mean value of 760 µm), they are usually longer than the diameter of the axes, and are much longer than the lowermost branchlet segment. The lower stipulodes are sometimes of unequal lengths, at 161–475 µm long, with a mean value of 293 µm. The branchlets are 9–12 in whorl and generally much shorter than the internodes, at 2.0–4.3 cm long. The branchlets of the uppermost 1–2 youngest whorls are even shorter, at just 0.2–2.0 cm long. The lowermost basal segments of the branchlets are ecorticated, and are very short at 208–479 µm long (mean value 343 µm) and 189–470 µm wide (mean value 318 µm). These segments are hidden behind the upper stipulodes (Figure 2A). The branchlets consist of 7–10 segments, with the lowermost segments always being ecorticated, followed by 4–6 corticated segments and 2–5 ecorticated distal segments with a tiny acute end cell on top, surrounded by a ring of bract cells (Figure 2C,F). The bract cells (5–8) are well developed (220–843 µm long), slender, and acute, and are shorter than the bracteoles. The two bracteoles are very long (990–1948 µm), at 1–2.5 × longer than the oogonia and oospores (Figure 2B). All of the fertile specimens are monoecious with conjoined gametangia (Figure 2B). Gametangia usually occur only at the nodes of corticated segments, and are rarely observed at the lowest nodes just above the ecorticated segment. The gametangia are mainly solitary, and very rarely geminate. The oogonia are elliptical to elongated oval in shape, are yellow or greenish in colour, and generally have constricted coronulae. The length of the oogonia (without coronula) is (600) 700–850 (900) µm, and the width of the oogonia is 417–575 (600) µm. The length of the coronula is 69–125 (150) µm, and the width of the coronula is (127) 160–200 (250) µm. The oospores are elliptical in shape and black in colour, with a length of (539) 600–685 µm, a width of 375–475 (500) µm, and 10–13 striae. The antheridiae are tetrascutate with a diameter of (300) 350–400 (450) µm. The dried specimens are stored at the herbarium of the University of Rostock (ROST).
2.3. Phylogenetic Analysis
The three individuals collected on Sardinia had identical rbcL and matK sequences. The BLAST of the GenBank nucleotide collection under default settings with rbcL from the Sardinian samples as query sequences matched the individuals to C. zeylanica from New Caledonia (AB440257) with 100% identity. One basepair (bp) substitution (99% identity) was detected for two further C. zeylanica (HQ380481: Sri Lanka, AY720934: Taiwan), but also for a sequence belonging to C. hydropitys Rchb. (HQ380464: Puerto Rico).
The BLAST of the GenBank nucleotide collection using matK from the Sardinian samples as query sequences matched the individuals with 99% identity (1 bp substitution) to C. zeylanica from Myanmar (MT739758). Chara guairensis R.M.T.Bicudo (KY656924) and C. hydropitys (KY656921) differed from the Sardinian samples by 15 bp substitutions (98% identity), respectively.
Phylogenetic analyses were performed for rbcL and matK separately to confirm the species identified through the BLAST search. The final rbcL alignment was trimmed to 1051 bp. Within the subsect. Willdenowia, 30 variable sites were identified. In the rbcL tree (Figure 3), the relationships within the subsect. Willdenowia were ambiguous, because several nodes did not have significant supports. The specimens from Sardinia formed a cluster together with C. zeylanica, but only with a low level of support (BS: 50%, PP: 0.6). The phylogeny based on the rbcL gene sequences only could not be resolved, and relationships of C. zeylanica to other species of subsect. Willdenowia were ambiguous.
The final matK alignment was trimmed to 970 bp. Within the subsect. Willdenowia, 64 variable sites were identified. Phylogenetic analysis of the matK alignment provided strong bootstrap support for the sequences from the Sardinian samples forming a monophyletic clade with C. zeylanica sequence: MT739758 (BS: 100% and BP: 1, Figure 4). The matK phylogeny assigned the Sardinian specimens to C. zeylanica, and differentiated them from other species of subsect. Willdenowia (C. guarensis, C. rusbyana M.Howe, C. haitensis Turpin, C. foliolosa Muhl. ex Willd.) and sect. Imahoria (C. hydropitys).
3. Discussion
3.1. Taxonomical Remarks
The specimens collected at Cala Fuili (Sardinia) were shown to qualify, based on their morphological characters, as a taxon belonging to subsect. Willdenowia because they are diplostephaneous with triplostichous cortication and have ecorticated basal segments of otherwise corticated branchlets [23]. Following the approach of van Raam [28], who distinguished 20 species within subsect. Willdenowia—in contrast to Wood [23], who identified a monospecific subsection—the question of to which species the specimens belong is discussed in detail below.
Van Raam [28] analyzed systematically the problem of gymnopodial (ecorticated first branchlet segment) taxa of the genus Chara L. using a stepwise approach. A total of 37 taxa of the genus Chara were found to share the character of an ecorticated basal branchlet segment. We recall that a taxon is a taxonomic unit of any rank, which can be species, but also varieties and forms. Eight taxa from subsect. Willdenowia can be excluded because they are haplostephaneous (and can thus be assigned to sect. Imahoria J. van Raam). Of the remaining 29 diplostephaneous taxa, C. kenoyeri M.Howe and C. rusbyana can be excluded here because they are dioecious. As a haplostichous species, Chara pseudohydropitys Imahori belongs to section Aghardia R.D.Wood, and can also be excluded here. Similarly, C. foliolosa, C. tenuifolia (Allen ex R.D.Wood) R.D.Wood, C. guairensis, C. haitensis, C. indica Bertero ex Spreng., C. martiana Wallman, and C. paucicorticata Cáceres can be excluded because they have octoscutate antheridia. Unlike the specimens described here, Chara drouetii (R.D.Wood) R.D.Wood, C. michauxii (A.Braun) Kütz., and C. formosa C.B.Rob. are characterised by a sejoined gametangia arrangement. Chara cubensis Allen, C. depauperata Allen, C. oerstediana A.Braun, and C. diaphana (Meyen) R.D.Wood have fewer than four corticated branchlet segments, whereas all the specimens found in Sardinia have at least four corticated segments. According to van Raam [28], the remaining 12 taxa belong to C. zeylanica as varieties or forms based on quantitative characters, such as spine length relative to axis diameter and the length of the stipulodes.
At this stage, we can conclude that the Sardinian specimens fit the character combination of C. zeylanica. Because van Raam [28] failed to provide an adequate description of infraspecific taxa beyond offering a series of tables, the specimens discussed here will not be related to varieties or forms. In any case, the specimens clearly do not belong to C. foliolosa, which can be found in the northernmost distribution range of subsect. Willdenowia in North America [25].
However, a sound comparison between our specimens and specimens described by other authors [39,40,43,54] is still impossible because of the different taxonomic concepts applied. Taking Wood [23] as a basis, many authors [27,45,55] did not take antheridia morphology into account. Thus, it is extremely difficult to compare their data with the recent concept proposed by van Raam [28].
To obtain independent proof of the morphology-based determination, phylogenetic analyses were performed with the regularly used barcode markers rbcL and matK. The analyses classified independently the individuals from Sardinia along with other C. zeylanica. Morphologically similar species such as C. foliolosa, C. haitensis, and C. rusbyana (previously considered to be varieties or forms of C. zeylanica sensu Wood) could be excluded through alignment with GenBank sequences. The phylogenetic analyses showed that C. hydropitys, a haplostephaneous species belonging to sect. Imahoria, is closely related to the abovementioned Willdenowia species, consistent with the findings of previous studies [43,56,57]. The phylogenetic relationships between C. zeylanica and C. hydropitis were not evident based on rbcL sequence data. However, the Sardinian samples were shown to have rbcL gene sequences identical to those of a C. zeylanica individual from GenBank (HQ380481), which made the categorisation unambiguous. The assignment of the specimens to this taxon was supported by the results of a matK analysis, which showed that C. zeylanica obtained from GenBank (MT739758) formed a monophyletic clade together with the Sardinian specimens [43,56,57]. Thus, the genetic classification based on the rbcL and matK sequences clearly supported the morphological determination of the individuals collected at Cala Fuili. The phylogeny of the subsect. Willdenowia was not the main focus of this study. Nevertheless, in order to test the phylogeny of Willdenowia species in future studies, the taxonomic and geographical basis for an analysis should be broadened, and additional molecular data should be gathered.
3.2. Status and Threats
Many charophyte species and their habitats are threatened throughout Europe, and are mentioned in several national Red Lists [4]. Sardinia has a key role to play in the conservation of Characeae in the Mediterranean region [30,58,59]. Becker [30] identified numerous Sardinian hotspots for the conservation of charophytes, and proposed specific action plans that mainly focused on Characeae in brackish habitats. The Sardinian site where C. zeylanica has been found is in the hotspot area between Orosei and Capo Comino.
In contrast to rare and threatened taxa, introduced non-native species can become invasive and cause ecological damage, as the example of Nitellopsis obtusa (Desv.) J.Groves in North America shows [60]. However, the examples of two alien charophyte species with mainly intertropical distribution that were previously introduced into Europe have so far not been found to have any serious environmental impacts. Both species, Chara fibrosa (including ssp. benthamii) and Chara c.f. chrysospora, were probably introduced by humans into rice fields in Southern France and Northern Italy through the importation of contaminated rice seeds [32,46,47,61]. Moreover, while the presence of a population of Chara fibrosa ssp. benthamii was recorded on the Greek island of Crete [48], it appears that it has been extinct since 2010 [62].
Chara zeylanica cannot currently be considered an invasive species among the European charophyte flora. For the moment, the Sardinian population is very small, and is limited to a single and relatively isolated location. Although the species has a high rate of fertility in Sardinia, strong dispersal cannot be expected at this stage. Nevertheless, the development of the Sardinian population of C. zeylanica should be monitored.
Although the abovementioned intertropical species Chara fibrosa and C. c.f. chrysospora were probably introduced into Europe by anthropogenic factors [47], this is unlikely to be the case for C. zeylanica. The Sardinian site is situated more than 100 km away from the nearest rice fields. The surrounding land is used primarily for grazing sheep and small-scale tourism. Other anthropogenic dispersal pathways (e.g., fishery, bathing, or diving) also appear to be unlikely. On the other hand, Sardinia is an important interim stop for birds migrating between Europe and Africa. As the nearest previous records of the presence of C. zeylanica are from a Saharan pond in Algeria at least 88 years ago [45 and literature therein], and from Senegal and Egypt [26,27], we assume that the species was introduced into Sardinia by migrating water birds. However, against the backdrop of climate change, future investigations of C. zeylanica and other Characeae should consider whether rice fields in Sardinia and throughout the Mediterranean area play a role in the dispersal of the species.
4. Material and Methods
4.1. Hydrochemical and Morphological Analyses
Hydrochemical analyses were conducted in a laboratory according to standard methods and national DIN norms, as published by Wasserchemische Gesellschaft [63,64,65]. The nutrient concentrations (NH4-N, NO3-N, total N, PO4-P and total P) were measured using a photometer (CADAS 200 by Dr Lange). The cation concentrations (Ca, Mg) were determined by means of an atomic absorption spectrometer (SpectrAA 55 by Varian). The pH values were analyzed using WTW Multi 3510 IDS. The conductivity, salinity, and chloride levels were determined using WTW Cond 3130, with the specific probe being applied in each case.
The morphological analysis was done by means of a stereo microscope (SZX16; Olympus, Tokyo, Japan) equipped with a digital camera for recording photographs.
4.2. DNA Barcoding
The total genomic DNA was extracted using the DNeasy Plant Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol. Partial sequences of the rbcL and matK genes were amplified using the primers rbcL-1a (5′-TCG TGT AAC TCC ACA ACC TG-3′) and rbcL-1b (5′-TAC TCG GTT AGC TAC AGC TC-3′), and matK-F2 (5′-GAA TGA GCT TAA ACA AGG ATT C-3′) and matK-R1b (5′-GCA GCC TTA TGA ATT GGA TAG C-3′). The PCR tests were performed in a 30 µL reaction volume with a Taq PCR Master Mix (Qiagen, Hilden, Germany) consisting of 2.5 mM MgCl2 (final concentration), and 0.5 pmol of each primer. The PCR products were extracted from agarose gels following the protocol of the Biometra-innuPrep Gel Extraction Kit (Analytik Jena, Jena, Germany), and were sequenced directly using a 3130×L Genetic Analyser (Applied Biosystems, New York, NY, USA) with sequencing primers identical to the primers that were used for the PCR reaction. The quality of the chromatograms of the generated sequences were checked using the BIOEDIT software [66]. The nucleotide sequences identified in this study have been deposited in the GenBank (MZ648319- MZ648324).
Sequences from three specimens collected in Sardinia were submitted to the National Center for Biotechnology Information’s (NCBI) Basic Local Alignment Search Tool (BLAST) [67] to allow them to be checked against the nucleotide collection in the GenBank in order to identify other Chara sequences with high scoring similarity pairs (HSP) in the NCBI web server. The phylogenetic analysis was performed with the sequence data from the Chara specimens collected in Sardinia, and with data on closely related taxa in the GenBank’s nucleotide database (https://www.ncbi.nlm.nih.gov/nuccore) for both the rbcL and matK sequences separately, because the sequences available in the GenBank were completely different. Alignments were created and trimmed using BIOEDIT software [66]. Identical sequences were merged into one entry. Sequences differing only in length were also reduced to one genotype. If different taxa had identical sequences, they were retained in the alignment (Table 1). The rbcL dataset contained 36 sequences belonging to nine species of the subsect. Willdenowia, and seven of haplostephanous species belonging to the sect. Charopsis, Protochara and Imahoria, and to the subsect. Wallmania and Agardhia. In addition, Nitellopsis obtusa was used as the outgroup (Table 1). For the matK dataset, the three Sardinien samples of C. zeylanica were analysed together with 13 sequences belonging to six species of the subsect. Willdenowia and Agardhia, and one species of the sect. Charopsis and Imahoria, respectivly. Nitellopsis obtusa was used as the outgroup (Table 1). Phylogenetic trees were created using the Maximum likelihood (ML) method and Bayesian inference (BI) analysis. The best-fit model of sequence evolution was determined using MEGA v. X [68]. The ML method was applied using MEGA v. X [68], with the HKY+G+I model used as the nucleotide substitution model for the rbcL dataset, and the GTR+G+I model used for the matK dataset. Branch supports were evaluated using 1000 bootstrap replicates (BS). MrBayes 3.2.7 [69] was used for the BI method. Two independent runs with four chains were run for 10 million generations using the MCMC method. Calculations of the consensus tree, including clade posterior probability (PP), were performed based on the trees sampled after the chains converged using Tracer 1.7 [70]. The first 25% were discarded as burn-in.
Author Contributions
Conceptualization, R.B., H.S. and P.N.; methodology, R.B., H.S. and P.N.; formal analysis, R.B., H.S. and P.N.; investigation, R.B.; writing—original draft preparation, R.B., H.S. and P.N.; writing—review and editing, R.B., H.S. and P.N.; visualization, R.B., H.S. and P.N. All authors have read and agreed to the published version of the manuscript.
Funding
Part of the equipment was funded by the European Regional Development Fund (ERDF, UHRO26).
Acknowledgments
We are very greatful to Melanie Willen (University of Oldenburg) for carrying out the hydrochemical analyses. The acquisition of some of the equipment was supported by a grant of the European Regional Development Fund (ERDF, UHRO26). We thank our anonymous reviewers and Angelo Troia, our scientific editor, for their helpful comments and suggestions, which greatly improved the final version of our paper. We thank Miriam Hils-Cosgrove for thorough language editing.
Conflicts of Interest
No potential conflict of interest was reported by the authors.
References
- Karol, K.G.; McCourt, R.M.; Cimino, M.T.; Delwiche, C.F. The closest living relatives of land plants. Science 2001, 294, 2351–2353. [Google Scholar] [CrossRef] [PubMed]
- Schubert, H.; Blindow, I.; Bueno, N.C.; Casanova, M.T.; Pełechaty, M.; Pukacz, A. Ecology of charophytes—Permanent pioneers and ecosystem engineers. Perspect. Phycol. 2018, 5, 61–74. [Google Scholar] [CrossRef]
- Doege, A.; van de Weyer, K.; Becker, R.; Schubert, H. Chapter 8 Bioindikation mit Characeen [Bioindication using Characeae]. In Armleuchteralgen. Die Characeen Deutschlands [Stoneworts. Characeae of Germany]; Arbeitsgruppe Characeen Deutschlands, Ed.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 97–138. [Google Scholar]
- Becker, R. Chapter 10 Gefährdung und Schutz von Characeen [Threats and conservation of charophytes]. In Armleuchteralgen. Die Characeen Deutschlands [Stoneworts. Characeae of Germany]; Arbeitsgruppe Characeen Deutschlands, Ed.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 149–191. [Google Scholar]
- Azzella, M. Italian volcanic lakes. A diversity hotspot and refuge for European charophytes. J. Limnol. 2014, 73, 502–510. [Google Scholar] [CrossRef] [Green Version]
- Blindow, I.; Garniel, A.; Munsterhjelm, R.; Nielsen, R. Conservation and threats. Proposal for a red data book for charophytes in the Baltic Sea. In Charophytes of the Baltic Sea; Schubert, H., Blindow, I., Eds.; Gantner, Ruggel: Königstein, Germany, 2003; pp. 251–260. [Google Scholar]
- Schwarzer, A. Aktionsplan Vielästige Glanzleuchteralge (Nitella hyalina (DC.) C. Agardh). [Action Plan for Many-Branched Stonewort (Nitella hyalina (DC.) C. Agardh)]; Baudirektion Kanton Zürich, Amt für Landschaft und Natur: Zürich, Switzerland, 2019. [Google Scholar]
- Blindow, I. Schwedische Artenschutzprogramme für bedrohte Characeen [Swedish conservation programmes for endangered Characeae]. Rostock. Meeresbiol. Beitr. 2008, 19, 23–28. [Google Scholar]
- Stewart, N. Important Stonewort Areas. An Assessment of the Best Areas for Stoneworts in the United Kingdom (Summary); Plantlife International: Salisbury, UK, 2004. [Google Scholar]
- Marquardt, R.; Schubert, H. Photosynthetic characterisation of Chara vulgaris in bioremediation ponds. Charophytes 2009, 2, 1–8. [Google Scholar]
- Braun, M.; Foissner, I.; Löhring, H.; Schubert, H.; Thiel, G. Characean algae: Still a valid model system to examine fundamental principles in plants. In Progress in Botany; Esser, K., Löttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 193–220. [Google Scholar]
- Selig, U.; Steinhardt, T.; Schubert, H. Interannual variability of submerged vegetation in a brackish coastal lagoon on the southern Baltic Sea. Ekológia 2009, 28, 412–423. [Google Scholar] [CrossRef]
- Jäger, D. Beiträge zur Characeen-Flora Vorarlbergs (Österreich) [Contributions to the Characeae Flora of Vorarlberg (Austria)]. Ber. Des Nat.-Med. Ver. Innsbr. 2000, 87, 67–85. [Google Scholar]
- Burns, N.M.; Rutherford, J.C.; Clayton, J.S. A monitoring and classification system for New Zealand lakes and reservoirs. Lake Reserv. Manag. 1999, 15, 255–271. [Google Scholar] [CrossRef]
- van Raam, J.C. Handboek Kranswieren; Chara Boek: Hilversum, The Netherlands, 1998. [Google Scholar]
- Gutowski, A.; Hofman, G.; Leukart, P.; Melzer, A.; Mollenhauer, M.; Schmedtje, U.; Schneider, S.; Tremp, H. Trophiekartierung von aufwuchs- und makrophytendominierten Fließgewässern. Trophic monitoring of emergent and macrophyte-dominated rivers and streams. Inf. Bayer. Landesamtes Wasserwirtsch. 1998, 4/98, 1–501. [Google Scholar]
- Melzer, A. Möglichkeiten einer Bioindikation durch submerse Makrophyten—Beispiele aus Bayern [Bioindication opportunities through submerged macrophytes—Examples from Bavaria]. Beiträge Angew. Gewässerökologie Norddtschl. 1994, 1, 92–102. [Google Scholar]
- Kohler, A. Wasserpflanzen als Belastungsindikatoren [Aquatic plants as stress indicators]. Dechen. Beih. 1982, 26, 31–42. [Google Scholar]
- Krause, W. Characeen als Bioindikatoren für den Gewässerzustand [Characeae as bioindicators of water quality]. Limnol.-Ecol. Manag. Inland Waters 1981, 13, 399–418. [Google Scholar]
- Migula, W. Die Characeen Deutschlands, Oesterreichs und der Schweiz [The Characeae of Germany, Austria and Switzerland]; Eduard Kummer: Leipzig, Germany, 1897. [Google Scholar]
- Deutschlands, A.C. (Ed.) Chapter 9: Die Armleuchteralgen-Gesellschaften Deutschlands. In Armleuchteralgen. Die Characeen Deutschlands [Stoneworts. Characeae of Germany]; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Korsch, H. The worldwide range of the Charophyte species native to Germany. Rostock. Meeresbiol. Beitr. 2018, 45–96. [Google Scholar]
- Wood, R.D.; Imahori, K. A revision of the Characeae. First Part: Monograph of the Characeae; Cramer: Weinheim, Germany, 1965. [Google Scholar]
- Wood, R.D. New combinations and taxa in the revision of Characeae. Taxon 1962, 11, 7–25. [Google Scholar] [CrossRef]
- Proctor, V.W.; Griffin, D.G.; Hotchkiss, A.T. A Synopsis of the Genus Chara, Series Gymnobasalia (Subsection Willdenowia RDW). Am. J. Bot. 1971, 58, 894. [Google Scholar] [CrossRef]
- Corillion, R.; Guerlesquin, M. Notes phytogéographiques sur les Charophycées d’Egypte [Flora geographical notes on the Charophyceae of Egypt]. Rev. Algol. 1971, 10, 177–191. [Google Scholar]
- Guerlesquin, M. Recherches sur Chara zeylanica Klein ex Willd. (Charophycées) d’Afrique occidentale [Research on Chara zeylanica Klein ex Willd. (Charophyceae) from West Africa]. Rev. Algol. 1971, 10, 231–247. [Google Scholar]
- van Raam, J.C. Annotated Bibliography of the Characeae, 2nd edition. CD.; Chara Boek: Hilversum, The Netherlands, 2010. [Google Scholar]
- van Raam, J.C. A matrix key for the determination of Characeae. Rostock. Meeresbiol. Beitr. 2009, 22, 53–55. [Google Scholar]
- Becker, R. The Characeae (Charales, Charophyceae) of Sardinia (Italy): Habitats, distribution and conservation. Webbia 2019, 74, 83–101. [Google Scholar] [CrossRef]
- Romanov, R.E.; Napolitano, T.; van de Weyer, K.; Troia, A. New records and observations to the Characean flora of Sicily (Italy). Webbia 2019, 74, 111–119. [Google Scholar] [CrossRef]
- Mouronval, J.-B.; Baudouin, S.; Borel, N.; Soulié-Märsche, I.; Klesczewski, M.; Grillas, P. Guide des Characées de France Méditerranéenne [Guide of Characeae of the French Mediterranean Region]; Office National de la Chasse et de la Faune Sauvage: Paris, France, 2015.
- Christia, C.; Tziortzis, I.; Fyttis, G.; Kashta, L.; Papastergiadou, E. A survey of the benthic aquatic flora in transitional water systems of Greece and Cyprus (Mediterranean Sea). Bot. Mar. 2011, 54, 169–178. [Google Scholar] [CrossRef]
- Bazzichelli, G.; Abdelahad, N. Alghe D’acqua Dolce d’Italia: Flora Analitica Delle Caroficee [Freshwater Algae of Italy: Analytical Flora of Charophytes]; Ministero dell’Ambiente e Della Tutela del Territorio e del Mare, Sapienza Università di Roma: Roma, Italy, 2009.
- Cirujano, S.; Cambra, J.; Sánchez-Castillo, P.M.; Meco, A.; Flor-Arnau, N. Flora Ibérica. Algas Continentales. Carófitos (Characeae); Real Jardin Botánico: Madrid, Spain, 2008. [Google Scholar]
- Blaženčić, Z.; Stevanovič, V.; Blaženčić, J.; Stevanovič, B. Red data list of charophytes in the Balkans. Biodivers. Conserv. 2006, 15, 3445–3457. [Google Scholar] [CrossRef]
- Duarte, T.; Almeida, T.S.; Albertoni, E.F.; Palma-Silva, C. Role of the propagule bank in reestablishing submerged macrophytes after removal of free-floating plants for recovery of a shallow lake in Southern Brazil. Ecol. Austral 2020, 30, 239–250. [Google Scholar] [CrossRef]
- Langangen, A. Some finds of Charophytes from East-Africa (Zambia, Tanzania, Kenya and Somalia). Acta Musei Natl. Pragae Ser. B Hist. Nat. 2015, 71, 229–248. [Google Scholar]
- Casanova, M.T. An overview of Chara L. in Australia (Characeae, Charophyta). Aust. Syst. Bot. 2005, 18, 25–39. [Google Scholar] [CrossRef]
- Leghari, S.M.; Langangen, A. Fresh water algae of Sindh, VI. Charales (Charophyta) from fresh and brackish water of Sindh, Pakistan. J. Biol. Sci. 2001, 1, 461–465. [Google Scholar] [CrossRef] [Green Version]
- Guerlesquin, M. Contribution a l’étude des Characées des Petites Antilles [Contribution to the research of Charophytes of the Lesser Antilles]. Rev. Hydrobiol. Trop. 1983, 16, 213–233. [Google Scholar]
- Zaneveld, J.S. The Charophyta of Malaysia and adjacent countries. Blumea 1940, 4, 1–223. [Google Scholar]
- Mjelde, M.; Swe, T.; Langangen, A.; Ballot, A. A contribution to the knowledge of charophytes in Myanmar; morphological and genetic identification and ecology notes. Bot. Lett. 2021, 168, 102–109. [Google Scholar] [CrossRef]
- Romanov, R.E.; Barinova, S.S. The charophytes of Israel: Historical and contemporary species richness, distribution and ecology. Biodivers. Res. Conserv. 2012, 25, 67–74. [Google Scholar] [CrossRef]
- Muller, S.D.; Rhazi, L.; Soulie-Märsche, I.; Benslama, M.; Bottollier-Curtet, M.; Daoud-Bouattour, A.; Belair, G.D.; Ghrabi-Gammar, Z.; Grillas, P.; Paradis, L.; et al. Diversity and distribution of Characeae in the Maghreb (Algeria, Morocco, Tunisia). Cryptogam. Algol. 2017, 38, 201–252. [Google Scholar] [CrossRef]
- Abdelahad, N.; Piccoli, F. Report on Charophytes from rice fields in northern Italy including the alien species Chara fibrosa ssp. benthamii. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2018, 152, 599–603. [Google Scholar] [CrossRef]
- Soulié-Märsche, I.; Triboit, F.; Despréaux, M.; Rey-Boissezon, A.; Laffont-Schwob, I.; Thiéry, A. Evidence of Chara fibrosa Agardh ex Bruzelius, an alien species in South France. Acta Bot. Gall. Bot. Lett. 2013, 160, 157–163. [Google Scholar] [CrossRef]
- Bergmeier, E.; Abrahamczyk, S. Current and historical diversity and new records of wetland plants in Crete, Greece. Willdenowia 2008, 38, 433–453. [Google Scholar] [CrossRef] [Green Version]
- Trbojević, I.; Marković, A.; Blaženčić, J.; Subakov Simić, G.; Nowak, P.; Ballot, A.; Schneider, S. Genetic and morphological variation in Chara contraria and a taxon morphologically resembling Chara connivens. Bot. Lett. 2020, 38, 1–14. [Google Scholar] [CrossRef]
- Langangen, A.; Ballot, A.; Nowak, P.; Schneider, S.C. Charophytes in warm springs on Svalbard (Spitsbergen): DNA barcoding identifies Chara aspera and Chara canescens with unusual morphological traits. Bot. Lett. 2019, 41, 1–8. [Google Scholar] [CrossRef]
- Nowak, P.; Schubert, H.; Schaible, R. Molecular evaluation of the validity of the morphological characters of three Swedish Chara sections: Chara, Grovesia, and Desvauxia (Charales, Charophyceae). Aquat. Bot. 2016, 134, 113–119. [Google Scholar] [CrossRef]
- Schneider, S.C.; Nowak, P.; Ammon, U.; von Ballot, A. Species differentiation in the genus Chara (Charophyceae): Considerable phenotypic plasticity occurs within homogenous genetic groups. Eur. J. Phycol. 2016, 51, 282–293. [Google Scholar] [CrossRef] [Green Version]
- Casanova, M.T.; Karol, K.G. A revision of Chara sect. Protochara, comb. et stat. nov. (Characeae: Charophyceae). Aust. Syst. Bot. 2014, 27, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Langangen, A.; Leghari, S.M. Some charophytes (Charales) from Pakistan. Stud. Bot. Hung. 2001, 32, 63–85. [Google Scholar]
- Casanova, M.T. Taxonomy of Daly River Macroalgae. In Recommendations for Nutrient Resource Condition Targets for the Daly River; Schult, J., Townsend, S., Douglas, M., Webster, I., Skinner, S., Casanova, M.T., Eds.; Charles Darwin University: Darwin, UK, 2008. [Google Scholar]
- Lee, E.-Y.; Kwang Chul, C.; Sang-Rae, L.; Kim, Y.H.; Lee, J.E.; Jee-Hwan, K. Re-examination of the morphological characteristics and rbcL gene sequences of Chara zeylanica Willdenow (Charales, Charophyceae) in Korea. Aquat. Bot. 2016, 134, 97–102. [Google Scholar] [CrossRef]
- Borges, F.R.; Necchi, O., Jr. Taxonomy and phylogeny of Chara (Charophyceae, Characeae) from Brazil with emphasis on the midwest and southeast regions. Phytotaxa 2017, 302, 101. [Google Scholar] [CrossRef]
- Casanova, M.T.; Becker, R. Lamprothamnium sardensis sp. nov.: A new species of Lamprothamnium for Europe (Characeae, Streptophyta). Phytotaxa 2021. submitted. [Google Scholar]
- Nowak, P.; van de Weyer, K.; Becker, R. The occurrence of sexual Chara canescens (Charales, Charophyceae) in Sardinia. Webbia 2019, 74, 103–109. [Google Scholar] [CrossRef]
- Pullmann, G.D.; Crawford, G. A Decade of Starry Stonewort in Michigan. Lakline Rep. Summer 2010, 2010, 36–42. [Google Scholar]
- Langangen, A. Chara fibrosa Agardh ex Bruzelius, a charophyte new to the European flora. Allionia 2000, 37, 249–252. [Google Scholar]
- Langangen, A. Charophytes (Charales) from Crete (Greece) collected in 2010. Fl. Medit. 2012, 22, 25–32. [Google Scholar] [CrossRef]
- Wasserchemische Gesellschaft in der GDCH, DIN (Ed.) DIN EN ISO 7980. Wasserbeschaffenheit [Water Quality]; Beuth-Verlag: Berlin, Germany, 2000. [Google Scholar]
- Wasserchemische Gesellschaft in der GDCH, DIN (Ed.) DIN EN ISO 38406. Deutsche Einheitsverfahren zur Wasser-, Abwasser- und Schlammuntersuchung, Kationen (Gruppe E) [German Standard Methods for the Examination of Water, Waste Water and Sludge—Cations (Group E)]; Beuth-Verlag: Berlin, Germany, 1993. [Google Scholar]
- Wasserchemische Gesellschaft in der GDCH, DIN (Ed.) DIN EN ISO 38405. Deutsche Einheitsverfahren zur Wasser-, Abwasser- und Schlammuntersuchung, Anionen (Gruppe D) [German Standard Methods for the Examination of Water, Waste Water and Sludge—Anions (Group D)]; Beuth-Verlag: Berlin, Germany, 1993. [Google Scholar]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Altschul, S. Basic local alignment search tool (BLAST). J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakayama, H.; Kasai, F.; Nozaki, H.; Watanabe, M.M.; Kawachi, M.; Shigyo, M.; Nishihiro, J.; Washitani, I.; Krienitz, L.; Ito, M. Taxonomic reexamination of Chara globularis (Charales, Charophyceae) from Japan based on oospore morphology and rbcL gene sequences, and the description of C. leptospora sp. nov. J. Phycol. 2009, 45, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Karol, K.G.; Alix, M.S.; Scribailo, R.W.; Skawinski, P.M.; Sleith, R.S.; Sardina, J.A.; Hall, J.D. New records of the rare North American endemic Chara brittonii (Characeae), with comments on its distribution. Brittonia 2018, 70, 277–288. [Google Scholar] [CrossRef]
- Kato, S.; Sakayama, H.; Sano, S.; Kasai, F.; Watanabe, M.M.; Tanaka, J.; Nozaki, H. Morphological variation and intraspecific phylogeny of the ubiquitous species Chara braunii (Charales, Charophyceae) in Japan. Phycologia 2008, 47, 191–202. [Google Scholar] [CrossRef]
- Hall, J.D.; Fučíková, K.; Lo, C.; Lewis, L.A. An assessment of proposed DNA barcodes in freshwater green algae. Cryptogam. Algol. 2010, 31, 529–555. [Google Scholar]
- McCourt, R.M.; Karol, K.G.; Casanova, M.T.; Feist, M. Monophyly of genera and species of Characeae based on rbcL sequences, with special reference to Australian and European Lychnothamnus barbatus (Characeae: Charophyceae). Aust. J. Bot. 1999, 47, 361–369. [Google Scholar] [CrossRef]
- Pérez, W.; Casanova, M.T.; Hall, J.D.; McCourt, R.M.; Karol, K.G. Phylogenetic congruence of ribosomal operon and plastid gene sequences for the Characeae with an emphasis on Tolypella (Characeae, Charophyceae). Phycologia 2016, 56, 230–237. [Google Scholar] [CrossRef]
- Sanders, E.R.; Karol, K.G.; McCourt, R.M. Occurrence of matK in a trnK groupII intron in charophyte green algae and phylogeny of the Characeae. Am. J. Bot. 2003, 90, 628–633. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Habitat photographs (left panels) and habitus (right panel) of specimens collected at Cala Fuili, Sardinia. Plants were collected in the widening of the small stream just in front of the bridge.
Figure 1.
Habitat photographs (left panels) and habitus (right panel) of specimens collected at Cala Fuili, Sardinia. Plants were collected in the widening of the small stream just in front of the bridge.

Figure 2.
Detailed photographs of C. zeylanica collected at Cala Fuili, Sardinia. (A) branchlet whorl with ecorticated basal segments; (B) conjoined gametangia; (C) ecorticated end segments with bract cells; (D) triplostichous main axis cortication with single spines; (E) diplostephanous stipulodes; (F) branchlet tip cell, surrounded by bract cells.
Figure 2.
Detailed photographs of C. zeylanica collected at Cala Fuili, Sardinia. (A) branchlet whorl with ecorticated basal segments; (B) conjoined gametangia; (C) ecorticated end segments with bract cells; (D) triplostichous main axis cortication with single spines; (E) diplostephanous stipulodes; (F) branchlet tip cell, surrounded by bract cells.

Figure 3.
Phylogenetic tree based on 1051 base pairs (bp) of the rbcL sequences. The evolutionary history was inferred using the GTR+G + I model. Maximum likelihood bootstrap values (>50%, ML, left) and Bayesian posterior probabilities (>0.5, BI, right) are given at the branches. The scale indicates sequence divergence in percent. The specimens collected in Sardinia are marked bold. Information about the section and the subsection of the species is given at the tree [24,29,53].
Figure 3.
Phylogenetic tree based on 1051 base pairs (bp) of the rbcL sequences. The evolutionary history was inferred using the GTR+G + I model. Maximum likelihood bootstrap values (>50%, ML, left) and Bayesian posterior probabilities (>0.5, BI, right) are given at the branches. The scale indicates sequence divergence in percent. The specimens collected in Sardinia are marked bold. Information about the section and the subsection of the species is given at the tree [24,29,53].

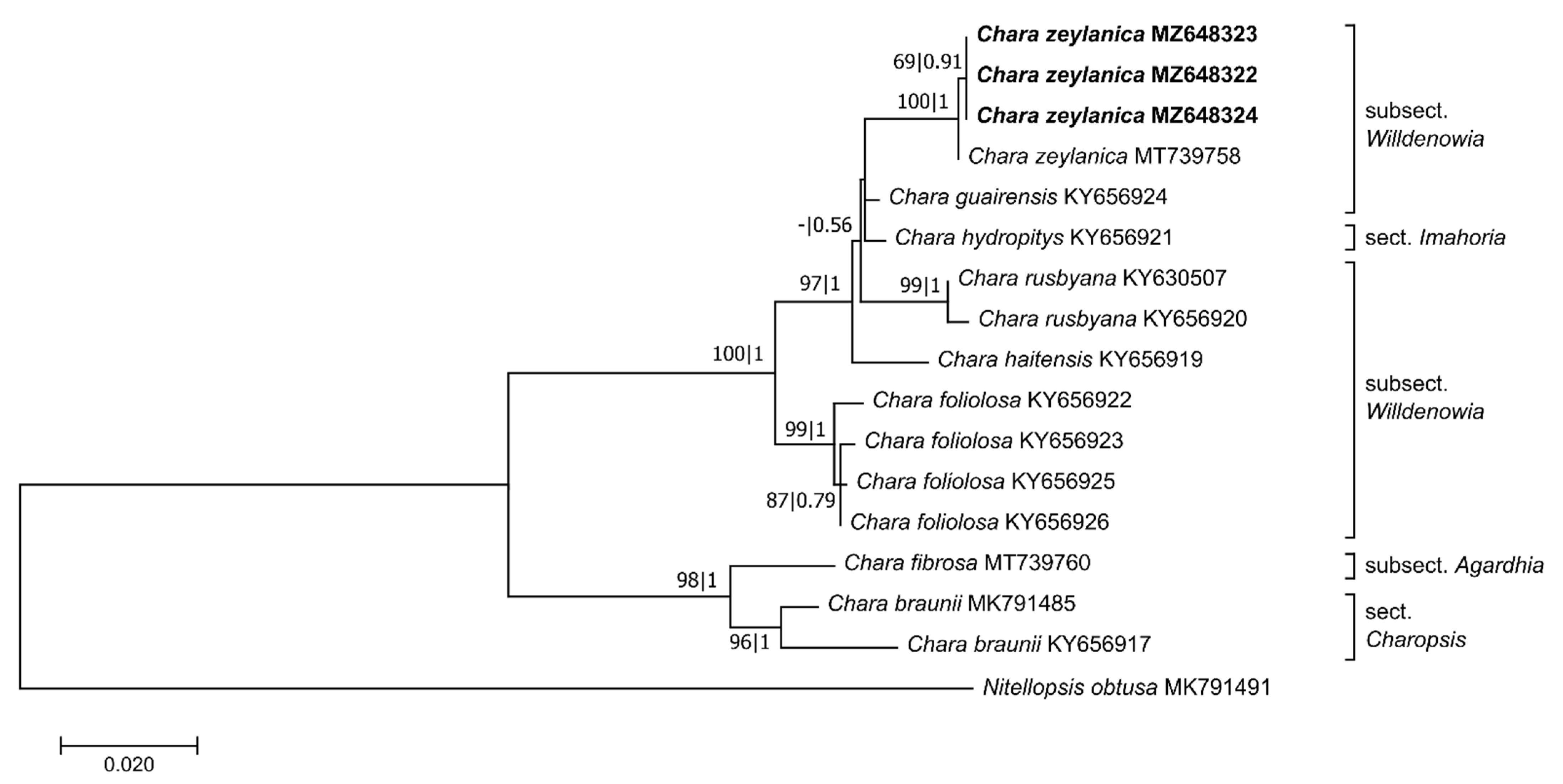
Figure 4.
Phylogenetic tree based on 970 base pairs (bp) of the matK sequences. The evolutionary history was inferred using the GTR+G+I model. Maximum likelihood bootstrap values (>50%, ML, left) and Bayesian posterior probabilities (>0.5, BI, right) are given at the branches. The scale indicates sequence divergence in percent. The specimens collected in Sardinia are marked bold. Information about the section and the subsection of the species is given at the tree [24,29,53].
Figure 4.
Phylogenetic tree based on 970 base pairs (bp) of the matK sequences. The evolutionary history was inferred using the GTR+G+I model. Maximum likelihood bootstrap values (>50%, ML, left) and Bayesian posterior probabilities (>0.5, BI, right) are given at the branches. The scale indicates sequence divergence in percent. The specimens collected in Sardinia are marked bold. Information about the section and the subsection of the species is given at the tree [24,29,53].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sample list of specimens used for phylogenetic analyses. Indicated are the accession numbers of the haplotypes (=non-redundant genotypes) downloaded from the GenBank. Information about the section and the subsection of the species is given in the third column [24,29,53] and identical sequences are given in the last column. n|a = not applicable.
Table 1.
Sample list of specimens used for phylogenetic analyses. Indicated are the accession numbers of the haplotypes (=non-redundant genotypes) downloaded from the GenBank. Information about the section and the subsection of the species is given in the third column [24,29,53] and identical sequences are given in the last column. n|a = not applicable.
| Marker | Species | Section/Subsection | Strain Designation and/or Collection Information | Accession | Reference | Redundant Accessions |
|---|---|---|---|---|---|---|
| rbcL | Chara australis | Protochara/- | S002/Unknown | AB440260 | [71] | - |
| Chara braunii | Charopsis/- | S036/Japan, Lake Ashino | AB440259 | [71] | - | |
| S019/Japan, Lake Haryu-numa | AB440258 | [71] | KJ395929 | |||
| GR10-UW37/Greece, Etoloakarnania | MK791476 | [59] | - | |||
| Chara brittonii | Grovesia/Willdenowia | KGK2610/USA, Wisconsin | MG880194 | [72] | MG880191, MG880186 | |
| KGK3120/USA, New Jersey | MG880206 | [72] | MG880205, MG880204, MG880203, MG880202, MG880201, MG880200, MG880199, MG880198, MG880197, MG880196, MG880195, MG880193, MG880192, MG880190, MG880189, MG880188, MG880187, MG880185, MG880184, MG880183 | |||
| Chara corallina | Protochara/- | SK026/Japan, Hiroshima | AB359167 | [73] | - | |
| Chara drouetii | Grovesia/Willdenowia | Proctor Loc 36/Guatemala, San Luis | HQ380445 | [74] | - | |
| KGK0467/Mexico, Quintana Roo | HQ380444 | [74] | - | |||
| Chara fibrosa | Agardhia/Agardhia | SK066/ Japan, Hiroshima | AB359168 | [73] | AB440261 | |
| Chara foliolosa | Grovesia/Willdenowia | Proctor 138/Mexico | MG880213 | [72] | HQ380448 | |
| NY 02146579/USA, Lake Erickson | MG880212 | [72] | MG880210, HQ380452, HQ380449 | |||
| NY 02145914/USA, Clopper Lake | MG880211 | [72] | MG880208, MG880207, KY656911, HQ380451, HQ380447, HQ380446 | |||
| NY 00739274/USA, Near New Deal | MG880209 | [72] | HQ380450 | |||
| SJRP31534/Brazil, Neves Paulista | KY656915 | [57] | KY656914, KY656907 | |||
| SJRP31929/Brazil, São Paulo | KY656912 | [57] | - | |||
| Chara guairensis | Grovesia/Willdenowia | SJRP31523/Brazil, São Paulo | KY656913 | [57] | - | |
| Chara haitensis | Grovesia/Willdenowia | SJRP28306/Brazil, Mato Grosso | KY656908 | [57] | - | |
| NY02145934/Michigan, USA | KX431011 | [75] | - | |||
| X-930/USA, Everglades | HQ380460 | [74] | HQ380459, HQ380458, HQ380457, HQ380456, HQ380455, HQ380454, HQ380453 | |||
| Chara hornemannii | Agardhia/Wallmania | NY00739162/Peru, Lima | KX431012 | [76] | - | |
| Chara hydropitys | Imahoria/- | SJRP28308/Brazil, Mato Grosso do Sul | KY656910 | [57] | - | |
| KGK0774/Puerto Rico, Lago Carite | HQ380464 | [74] | HQ380463, HQ380462, HQ380461 | |||
| Chara kenoyeri | Grovesia/Willdenowia | TP118/Panama, Gatun Lake | HQ380465 | [74] | - | |
| Chara longifolia | Agardhia/Wallmania | MB/Canada, Saskatchewan | AY170452 | [77] | - | |
| Chara martiana | Grovesia/Willdenowia | Proctor X-952/Venezuela, Caracas | HQ380467 | [74] | - | |
| Proctor TP097/Brazil, São Paulo | HQ380466 | [74] | - | |||
| Chara rusbyana | Grovesia/Willdenowia | SJRP28307/Brazil, Mato Grosso do Sul | KY656909 | [57] | KY630506 | |
| LG/ Unknown | AF097169 | [75] | AF097168 | |||
| Chara zeylanica | Grovesia/Willdenowia | n|a/Taiwan, Gueishan Island | AY720934 | unpubl. | - | |
| n|a/Australia: Elizabeth Creek | DQ076299 | unpubl. | - | |||
| S111/New Caledonia | AB440257 | [71] | KT343914, KT343913, AB359169, HQ380480, HQ380479, HQ380477, HQ380475, HQ380474, HQ380473, HQ380472, HQ380471, HQ380469, HQ380468 | |||
| Proctor X-574/Sri Lanka, Ceylon | HQ380481 | [74] | HQ380478, HQ380476, HQ380470 | |||
| RB-CZ119A | MZ648319 | this study | - | |||
| RB-CZ119B | MZ648320 | this study | - | |||
| RB-CT1119-B | MZ648321 | this study | - | |||
| Nitellopsis obtusa | S042/Germany, Lake Nehmitz | AB440263 | MK791482 | |||
| matK | Chara braunii | Charopsis/- | GR10-UW37/ Greece, Etoloakarnania | MK791485 | [59] | - |
| 48/ Brazil, São Paulo | KY656917 | [57] | - | |||
| Chara fibrosa | Agardhia/Agardhia | MY-33/Myanmar, Yezin | MT739760 | [43] | MT739765, MT739766, MT739768 | |
| Chara foliolosa | Grovesia/Willdenowia | SJRP31527/Brazil, Paulicéia | KY656925 | [57] | - | |
| SJRP31929/Brazil, São Paulo | KY656923 | [57] | - | |||
| SJRP31534/Brazil, Neves Paulista | KY656926 | [57] | - | |||
| SJRP28309/Brazil, Mato Grosso do Sul | KY656922 | [57] | - | |||
| Chara guairensis | Grovesia/Willdenowia | SJRP31523/Brazil, São Paulo | KY656924 | [57] | - | |
| Chara haitensis | Grovesia/Willdenowia | SJRP28306/Brazil, Mato Grosso | KY656919 | [57] | - | |
| Chara hydropitys | Imahoria/- | SJRP28308/Brazil, Mato Grosso do Sul | KY656921 | [57] | - | |
| Chara rusbyana | Grovesia/Willdenowia | n|a/Brazil. Mato Grosso do Sul | KY630507 | [57] | KY656916 | |
| SJRP28307/Brazil. Mato Grosso do Sul | KY656920 | [57] | - | |||
| Chara zeylanica | Grovesia/Willdenowia | MMYA-1/Myanmar, Inlay Lake | MT739758 | [43] | MT739759, MT739761, MT739762, MT739763, MT739764, MT739767 | |
| RB-CZ119A | MZ648322 | this study | - | |||
| RB-CZ119B | MZ648323 | this study | - | |||
| RB-CT1119-B | MZ648324 | this study | - | |||
| Nitellopsis obtusa | GEC4-1/Poland, Lake Lagowskie | MK791491 | [59] | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Becker, R.; Schubert, H.; Nowak, P. Chara zeylanica J.G.Klein ex Willd. (Charophyceae, Charales, Characeae): First European Record from the Island of Sardinia, Italy. Plants 2021, 10, 2069. https://doi.org/10.3390/plants10102069
AMA Style
Becker R, Schubert H, Nowak P. Chara zeylanica J.G.Klein ex Willd. (Charophyceae, Charales, Characeae): First European Record from the Island of Sardinia, Italy. Plants. 2021; 10(10):2069. https://doi.org/10.3390/plants10102069
Chicago/Turabian StyleBecker, Ralf, Hendrik Schubert, and Petra Nowak. 2021. "Chara zeylanica J.G.Klein ex Willd. (Charophyceae, Charales, Characeae): First European Record from the Island of Sardinia, Italy" Plants 10, no. 10: 2069. https://doi.org/10.3390/plants10102069
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.