
Exploitation and Conservation Status of the Thornback Ray (Raja clavata) in the Balearic Islands (Western Mediterranean)
 , ,
, ,  , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Stock Assessment for Exploitation Status
- Relative biomass (B/BMSY): the ratio between the current biomass (B) and the biomass that would provide the maximum sustainable yield (BMSY).
- Exploitation rate (F/FMSY): the ratio between the current fishing mortality (F), and the fishing mortality associated with the Maximum Sustainable Yield (FMSY) [33].
2.2. Genetic Analyses for Conservation Status
- R. clavata populations (N= 7): COI sequences data from different populations of R. clavata from the Mediterranean Sea and the Atlantic Ocean were obtained from GenBank. Only populations with 20 or more sequences from different individuals were kept (Supplementary Table S1).
- Rajiformes species (N= 42): this dataset was obtained by selecting the Rajiformes species from the Elasmobranchii dataset, once it was prepared (see below).
- Elasmobranchii species (N= 258): COI sequences from Elasmobranchii species with more than 15 sequences were downloaded from BOLDSystem v4 [45]. Only sequences with associated complete species names were included (i.e., sequences with abbreviations as sp. cf. var. in the species name were not considered). Sequences from complete mitochondrial genome projects and duplicate sequences were not included and IUPAC ambiguity codes were masked to N.
3. Results
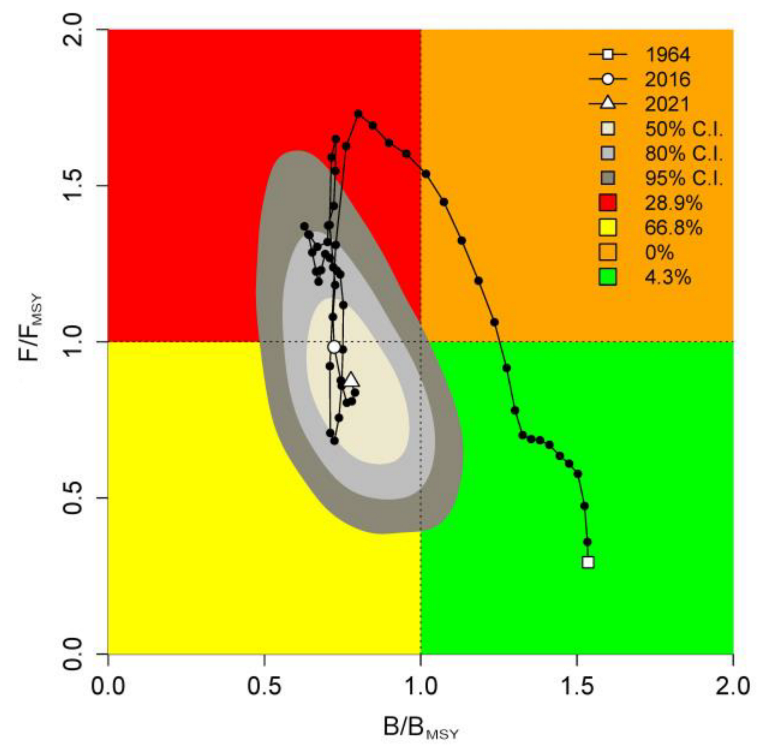
3.1. Exploitation Status
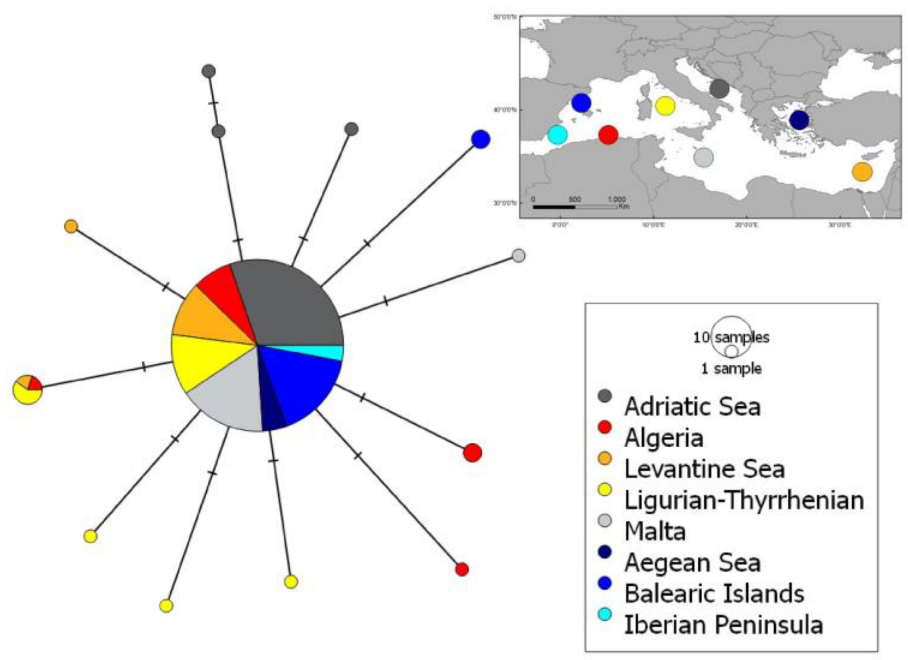
3.2. Conservation Status
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Browman, H.I.; Stergiou, K.I. Marine Protected Areas as a Central Element of Ecosystem-Based Management: Defining Their Location, Size and Number. Mar. Ecol. Prog. Ser. 2004, 274, 202–269. [Google Scholar] [CrossRef] [Green Version]
- Pikitch, E.K.; Santora, C.; Babcock, E.A.; Bakun, A.; Bonfil, R.; Conover, D.O.; Dayton, P.; Doukakis, P.; Fluharty, D.; Heneman, B.; et al. Ecosystem-Based Fishery Management. Science 2004, 305, 346–347. [Google Scholar] [CrossRef]
- Stevens, J.D.; Bonfil, R.; Dulvy, N.K.; Walker, P.A. The Effects of Fishing on Sharks, Rays, and Chimaeras (Chondrichthyans), and the Implications for Marine Ecosystems. ICES J. Mar. Sci. 2000, 57, 476–494. [Google Scholar] [CrossRef]
- Wetherbee, B.M.; Cortés, E. Food Consumption and Feeding Habits. In Biology of Sharks and Their Relatives; CRC press: Boca Raton, FL, USA, 2004; pp. 223–244. [Google Scholar]
- Ebert, D.; Stehmann, M. Sharks, Batoids, and Chimaeras of the North Atlantic. FAO Species Catalogue for Fishery Purposes. No. 7; FAO: Rome, Italy, 2013; ISBN 978-92-5-107466-4. [Google Scholar]
- Serena, F. Field Identification Guide to the Sharks and Rays of the Mediterranean and Black Sea. FAO Species identification guide for Fishery Purposes; Food and Agriculture Organization: Rome, Italy, 2005; p. 97. [Google Scholar]
- Bottari, T.; Rinelli, P.; Bianchini, M.L.; Ragonese, S. Stock Identification of Raja clavata L. (Chondrichthyes, Rajidae) in Two Contiguous Areas of the Mediterranean. Hydrobiologia 2013, 703, 215–224. [Google Scholar] [CrossRef]
- Follesa, M.C.; Marongiu, M.F.; Zupa, W.; Bellodi, A.; Cau, A.; Cannas, R.; Colloca, F.; Djurovic, M.; Isajlovic, I.; Jadaud, A.; et al. Spatial Variability of Chondrichthyes in the Northern Mediterranean. Sci. Mar. 2019, 83, 81–100. [Google Scholar] [CrossRef] [Green Version]
- Kadri, H.; Marouani, S.; Saïdi, B.; Bradai, M.N.; Bouaïn, A.; Morize, E. Age, Growth, Sexual Maturity and Reproduction of the Thornback Ray, Raja clavata (L.), of the Gulf of Gabès (South-Central Mediterranean Sea). Mar. Biol. Res. 2014, 10, 416–425. [Google Scholar] [CrossRef]
- Carbonara, P.; Bellodi, A.; Palmisano, M.; Mulas, A.; Porcu, C.; Zupa, W.; Donnaloia, M.; Carlucci, R.; Sion, L.; Follesa, M.C. Growth and Age Validation of the Thornback Ray (Raja clavata Linnaeus, 1758) in the South Adriatic Sea (Central Mediterranean). Front. Mar. Sci. 2020, 7, 586094. [Google Scholar] [CrossRef]
- Ramírez-Amaro, S.; Ordines, F.; Esteban, A.; García, C.; Guijarro, B.; Salmerón, F.; Terrasa, B.; Massutí, E. The Diversity of Recent Trends for Chondrichthyans in the Mediterranean Reflects Fishing Exploitation and a Potential Evolutionary Pressure towards Early Maturation. Sci. Rep. 2020, 10, 547. [Google Scholar] [CrossRef] [Green Version]
- Saglam, H.; Orhan, A. Reproductive Biology of Raja Clavata (Elasmobranchii: Rajidae) from Southern Black Sea Coast around Turkey. Helgol. Mar. Res. 2012, 66, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.R.; Shackley, S.E. Observations on Egg-Laying in the Thornback Ray. J. Fish Biol. 1995, 46, 903–904. [Google Scholar] [CrossRef]
- Capapé, C.; Guélorget, O.; Vergne, Y.; Quignard, J.P. Reproductive Biology of the Thornback Ray Raja clavata (Chondrichthyes: Rajidae) from the Coast of Languedoc (Southern France, Northern Mediterranean). Vie et Milieu 2007, 57, 83–90. [Google Scholar]
- Serra-Pereira, B.; Figueiredo, I.; Gordo, L.S. Maturation, Fecundity, and Spawning Strategy of the Thornback Ray, Raja Clavata: Do Reproductive Characteristics Vary Regionally? Mar. Biol. 2011, 158, 2187–2197. [Google Scholar] [CrossRef]
- Krstulović Šifner, S.; Vrgoč, N.; Dadić, V.; Isajlović, I.; Peharda, M.; Piccinetti, C. Long-Term Changes in Distribution and Demographic Composition of Thornback Ray, Raja clavata, in the Northern and Central Adriatic Sea. J. Appl. Ichthyol. 2009, 25, 40–46. [Google Scholar] [CrossRef]
- Guijarro, B.; Quetglas, A.; Moranta, J.; Ordines, F.; Valls, M.; González, N.; Massutí, E. Inter- and Intra-Annual Trends and Status Indicators of Nektobenthic Elasmobranchs off the Balearic Islands (Northwestern Mediterranean). Sci. Mar. 2012, 76, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Serena, F.; Mancusi, C.; Barone, M. Guida Pratica Di Identificazione Delle Razze (Rajidae) Del Mar Mediterraneo. Linee Guida per La Raccolte e l’analisis dei dati. Field Identification Guide to the Skates (Rajidae) of the Mediterranean Sea. Guidelines for Data Collection and Analysis. Biol. Mar. Mediterr. 2010, 17 (Suppl. 2), 204. [Google Scholar]
- Ellis, J. Raja clavata. The IUCN Red List of Threatened Species; 2016. [Google Scholar]
- Poisson, F. Review of the Available Data on Four Elasmobranches Species Caught by the French Fleets on the Mediterranean Coast (Gulf of Lions and Corsica). In Proceedings of the Workshop on Stock Assessment of Selected Species of Elasmobranchs in the GFCM area, Brussels, Belgium, 12–16 December 2011. [Google Scholar]
- Garofalo, G.; Gristina, M.; Fiorentino, F.; Cigala Fulgosi, F.; Norrito, G.; Sinacori, G. Distributional Pattern of Rays (Pisces, Rajidae) in the Strait of Sicily in Relation to Fishing Pressure. Hydrobiologia 2003, 503, 245–250. [Google Scholar] [CrossRef]
- Falsone, F.; Gancitano, V.; Geraci, M.L.; Sardo, G.; Scannella, D.; Serena, F.; Vitale, S.; Fiorentino, F. Assessing the Stock Dynamics of Elasmobranchii off the Southern Coast of Sicily by Using Trawl Survey Data. Fishes 2022, 7, 136. [Google Scholar] [CrossRef]
- Proceedings of the General Fisheries Commission for the Mediterranean Report of the Fourteenth Session of the Scientific Advisory Committee; Scientific Advisory Committee: Sofia, Bulgaria.
- Tsikliras, A.; Froese, R. Maximum Sustainable Yield. In Encyclopedia of Ecology; Fath, B., Ed.; Elsevier: Oxford, UK, 2019; Volume 1, pp. 108–115. [Google Scholar]
- Froese, R.; Winker, H.; Coro, G.; Palomares, M.L.D.; Tsikliras, A.C.; Dimarchopouplou, D.; Touloumis, K.; Demirel, N.; Scarcella, G.; de Souza Vianna, G.M.; et al. Catch Time Series as the Basis for Fish Stock Assessments: CMSY++ and Its Applications.; GEOMAR: Kiel, Germany, 2021. [Google Scholar]
- Petit-Marty, N.; Vázquez-Luis, M.; Hendriks, I.E. Use of the Nucleotide Diversity in COI Mitochondrial Gene as an Early Diagnostic of Conservation Status of Animal Species. Conserv. Lett. 2021, 14, e12756. [Google Scholar] [CrossRef]
- Spielman, D.; Brook, B.W.; Frankham, R. Most Species Are Not Driven to Extinction before Genetic Factors Impact Them. Proc. Natl. Acad. Sci. USA 2004, 101, 15261–15264. [Google Scholar] [CrossRef] [Green Version]
- Conover, D.O.; Clarke, L.M.; Munch, S.B.; Wagner, G.N. Spatial and Temporal Scales of Adaptive Divergence in Marine Fishes and the Implications for Conservation. J. Fish Biol. 2006, 69, 21–47. [Google Scholar] [CrossRef]
- Allendorf, F.W.; England, P.R.; Luikart, G.; Ritchie, P.A.; Ryman, N. Genetic Effects of Harvest on Wild Animal Populations. Trends Ecol. Evol. 2008, 23, 327–337. [Google Scholar] [CrossRef]
- Hare, M.; Nunney, L.; Schwartz, M.K.; Ruzzante, D.; Burford, M.; Waples, R.S.; Ruegg, K.; Palstra, F. Understanding and Estimating Effective Population Size for Practical Application in Marine Species Management. Conserv. Biol. 2011, 25, 438–449. [Google Scholar] [CrossRef] [Green Version]
- Pinsky, M.L.; Palumbi, S.R. Meta-Analysis Reveals Lower Genetic Diversity in Overfished Populations. Mol. Ecol. 2014, 23, 29–39. [Google Scholar] [CrossRef]
- Bernatchez, L.; Wellenreuther, M.; Araneda, C.; Ashton, D.T.; Barth, J.M.I.; Beacham, T.D.; Maes, G.E.; Martinsohn, J.T.; Miller, K.M.; Naish, K.A.; et al. Harnessing the Power of Genomics to Secure the Future of Seafood. Trends Ecol. Evol. 2017, 32, 665–680. [Google Scholar] [CrossRef] [Green Version]
- Gandra, M.; Assis, J.; Martins, M.R.; Abecasis, D. Reduced Global Genetic Differentiation of Exploited Marine Fish Species. Mol. Biol. Evol. 2021, 38, 1402–1412. [Google Scholar] [CrossRef]
- Petit-Marty, N.; Liu, M.; Tan, I.Z.; Chung, A.; Terrasa, B.; Guijarro, B.; Ordines, F.; Ramírez-Amaro, S.; Massutí, E.; Schunter, C. Declining Population Sizes and Loss of Genetic Diversity in Commercial Fishes: A Simple Method for a First Diagnostic. Front. Mar. Sci. 2022, 9, 872537. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. FishBase: A Global Information System on Fishes. Available online: https://www.fishbase.se/summary/Raja-clavata.html (accessed on 9 November 2022).
- Froese, R.; Winker, H.; Coro, G.; Demirel, N.; Tsikliras, A.C.; Dimarchopoulou, D.; Scarcella, G.; Quaas, M.; Matz-Lück, N. Status and Rebuilding of European Fisheries. Mar. Policy 2018, 93, 159–170. [Google Scholar] [CrossRef]
- Bertrand, J.A.; Gil de Sola, L.; Papaconstantinou, C.; Relini, G.; Souplet, A. The General Specifications of the MEDITS Surveys. Sci. Mar. 2002, 66, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Massuti, E.; Reñones, O. Demersal Resource Assemblages in the Trawl Fishing Grounds off the Balearic Islands (Western Mediterranean). Sci. Mar. 2005, 69, 167–181. [Google Scholar] [CrossRef] [Green Version]
- Spedicato, M.T.; Massutí, E.; Mérigot, B.; Tserpes, G.; Jadaud, A.; Relini, G. The MEDITS Trawl Survey Specifications in an Ecosystem Approach to Fishery Management. Sci. Mar. 2019, 83, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Anonymous. MEDITS Handbook, ninth version; MEDITS Working Group, 2017; p. 106. [Google Scholar]
- Ivanova, N.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D.N. Universal Primer Cocktails for Fish DNA Barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Ramírez-Amaro, S.; Ordines, F.; Picornell, A.; Castro, J.A.; Ramon, C.; Massutí, E.; Terrasa, B. The Evolutionary History of Mediterranean Batoidea (Chondrichthyes: Neoselachii). Zool. Scr. 2018, 47, 686–698. [Google Scholar] [CrossRef]
- Nei, M. Genetic Distance between Populations. In Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987; pp. 208–253. [Google Scholar]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-Based Registry for All Animal Species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE v5 Enables Improved Estimates of Phylogenetic Tree Confidence by Ensemble Bootstrapping. bioRxiv 2021. bioRxiv:2021.06.20.449169. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP v6: DNA Sequence Polymorphism Analysis of Large Datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Bandelt, H.; Forster, P.; Röhl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Ovenden, J.R.; Berry, O.; Welch, D.J.; Buckworth, R.C.; Dichmont, C.M. Ocean’s Eleven: A Critical Evaluation of the Role of Population, Evolutionary and Molecular Genetics in the Management of Wild Fisheries. Fish Fish. 2015, 16, 125–159. [Google Scholar] [CrossRef]
- Ramírez-Amaro, S.; Bassitta, M.; Picornell, A.; Ramon, C.; Terrasa, B. Environmental DNA: State-of-the-Art of Its Application for Fisheries Assessment in Marine Environments. Front. Mar. Sci. 2022, 9, 1004674. [Google Scholar] [CrossRef]
- Marongiu, M.F.; Porcu, C.; Bellodi, A.; Cannas, R.; Cau, A.; Cuccu, D.; Mulas, A.; Follesa, M.C. Temporal Dynamics of Demersal Chondrichthyan Species in the Central Western Mediterranean Sea: The Case Study in Sardinia Island. Fish Res. 2017, 193, 81–94. [Google Scholar] [CrossRef]
- FAO. Proceedings of the General Fisheries Commission for the Mediterranean-Report of the Twenty-Third Session of the Scientific Advisory Committee on Fisheries; FAO Fisheries and Aquaculture Report No. 1395; Scientific Advisory Committee on Fisheries: Rome, Italy. [CrossRef]
- Quetglas, A.; Guijarro, B.; Ordines, F.; Massutí, E. Stock Boundaries for Fisheries Assessment and Management in the Mediterranean: The Balearic Islands as a Case Study. Sci. Mar. 2012, 76, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Ordines, F.; Massutí, E.; Moranta, J.; Quetglas, A.; Guijarro, B.; Fliti, K. Balearic Islands vs Algeria: Two Nearby Western Mediterranean Elasmobranch Assemblages with Different Oceanographic Scenarios and Fishing Histories. Sci. Mar. 2011, 75, 707–717. [Google Scholar] [CrossRef]
- FAO. Proceedings of the General Fisheries Commission for the Mediterranean-Report of the sixteenth session of the Scientific Advisory Committee; FAO Fisheries and Aquaculture Report No. R1102; Scientific Advisory Committee on Fisheries: Rome, Italy; p. 251.
- Catalano, G.; Crobe, V.; Ferrari, A.; Baino, R.; Massi, D.; Titone, A.; Mancusi, C.; Serena, F.; Cannas, R.; Carugati, L.; et al. Strongly Structured Populations and Reproductive Habitat Fragmentation Increase the Vulnerability of the Mediterranean Starry Ray Raja asterias (Elasmobranchii, Rajidae). Aquat. Conserv. 2022, 32, 66–84. [Google Scholar] [CrossRef]
- Chevolot, M.; Hoarau, G.; Rijnsdorp, A.D.; Stam, W.T.; Olsen, J.L. Phylogeography and Population Structure of Thornback Rays (Raja clavata L., Rajidae). Mol. Ecol. 2006, 15, 3693–3705. [Google Scholar] [CrossRef]
- Pasolini, P.; Ragazzini, C.; Zaccaro, Z.; Cariani, A.; Ferrara, G.; Gonzalez, E.G.; Landi, M.; Milano, I.; Stagioni, M.; Guarniero, I.; et al. Quaternary Geographical Sibling Speciation and Population Structuring in the Eastern Atlantic Skates (Suborder Rajoidea) Raja clavata and R. straeleni. Mar. Biol. 2011, 158, 2173–2186. [Google Scholar] [CrossRef] [Green Version]
- Ballard, J.W.O.; Whitlock, M.C. The Incomplete Natural History of Mitochondria. Mol. Ecol. 2004, 13, 729–744. [Google Scholar] [CrossRef] [Green Version]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002; ISBN 0521639859. [Google Scholar]
- Nei, M.; Maruyama, T.; Chakraborty, R. The Bottleneck Effect and Genetic Variability in Populations. Evolution 1975, 29, 1–10. [Google Scholar] [CrossRef]
- Kenchington, E.; Heino, M.; Nielsen, E.E. Managing Marine Genetic Diversity: Time for Action? ICES J. Mar. Sci. 2003, 60, 1172–1176. [Google Scholar] [CrossRef] [Green Version]
- Oliver, F. Catálogo de La Fauna Marítima de Mallorca; Publicaciones del Instituto Social de la Marina: Palma, Spain, 1944. [Google Scholar]
- Maurin, C. Ecologie Ichthyologique Des Fonds Chalutables Atlantiques (de La Baie Ibéro-Marocaine à La Mauritanie) et de La Méditerranée Occidentale. Rev. Des Trav. De L’institut Des Pêches Marit. 1968, 32, 1–144. [Google Scholar]
- Barceló, F. Catálogo Metódico de Los Peces Que Habitan o Frecuentan Las Costas de Las Islas Baleares. Rev. De Los Prog. De Las Cienc. Exactas Físicas Y Nat. 1868, 18, 1–46. [Google Scholar]
- Lozano Rey, L. Ictiología Ibérica (Fauna Ibérica). Peces (Generalidades, Ciclóstomos y Elasmobranquios). Mus. Nac. De Cienc. Nat. 1928, 1, 1–692. [Google Scholar]
- Farriols, M.T.; Ordines, F.; Somerfield, P.J.; Pasqual, C.; Hidalgo, M.; Guijarro, B.; Massutí, E. Bottom Trawl Impacts on Mediterranean Demersal Fish Diversity: Not so Obvious or Are We Too Late? Cont. Shelf. Res. 2017, 137, 84–102. [Google Scholar] [CrossRef] [Green Version]
- Grau, A.M.; Mayol, J.; Oliver, J.; Riera, F.; Riera, I. Llibre Vermell Dels Peixos de Les Illes Balears; Conselleria de Medi Ambient, Agricultura i Pesca: Palma, Spain, 2015. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographical Area | N | NH | h ± SD | π ± SD |
|---|---|---|---|---|
| Balearic Islands | 31 | 2 | 0.125 ± 0.077 | 0.0002 ± 0.0004 |
| Ligurian–Tyrrhenian Sea | 26 | 5 | 0.406 ± 0.116 | 0.0008 ± 0.0008 |
| Adriatic Sea | 56 | 4 | 0.105 ± 0.056 | 0.0003 ± 0.0004 |
| Levantine Sea | 20 | 3 | 0.195 ± 0.115 | 0.0004 ± 0.0005 |
| Malta | 30 | 2 | 0.067 ± 0.061 | 0.0001 ± 0.0003 |
| Mediterranean Sea | 163 | 11 | 0.164 ± 0.040 | 0.0003 ± 0.0005 |
| Atlantic Ocean | 31 | 3 | 0.471 ± 0.074 | 0.0038 ± 0.0024 |
| Worldwide | 194 | 13 | 0.230 ± 0.040 | 0.0011 ± 0.0010 |
| Balearic Islands | Ligurian-Tyrrhenian Sea | Adriatic Sea | Levantine Sea | Malta | |
|---|---|---|---|---|---|
| Balearic Islands | X2 = 9.286 p = 0.098 | X2 = 5.276 p = 0.260 | X2 = 4.407 p = 0.221 | X2 = 2.984 p = 0.225 | |
| Ligurian–Tyrrhenian Sea | X2 = 9.286 p = 0.098 | X2 = 14.942 p = 0.037 * | X2 = 4.397 p = 0.494 | X2 = 8.410 p = 0.135 | |
| Adriatic Sea | X2 = 5.276 p = 0.260 | X2 = 14.942 p = 0.037 * | X2 = 6.705 p = 0.244 | X2 = 3.482 p = 0.481 | |
| Levantine Sea | X2 = 4.407 p = 0.221 | X2 = 4.397 p = 0.493 | X2 = 6.705 p = 0.244 | X2 = 3.723 p = 0.293 | |
| Malta | X2 = 2.984 p = 0.225 | X2 = 8.410 p = 0.135 | X2 = 3.482 p = 0.481 | X2 = 3.723 p = 0.293 |
| Dataset | Mean (95% CI) | Median (95% CI) |
|---|---|---|
| Mediterranean | 0.00034 (0.0003–0.0004) | 0.00036 (0.0002–0.0004) |
| Global | 0.0012 (0.0011–0.0013) | 0.00115 (0.0010–0.0012) |
| Rajiformes | 0.0040 (0.0032–0.0069) | 0.0030 (0.0022–0.0032) |
| Elasmobranchii | 0.0070 (0.0054–0.0077) | 0.0030 (0.0024–0.0033) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferragut-Perello, F.; Ramírez-Amaro, S.; Tsikliras, A.C.; Petit-Marty, N.; Dimarchopoulou, D.; Massutí, E.; Serrat, A.; Ordines, F. Exploitation and Conservation Status of the Thornback Ray (Raja clavata) in the Balearic Islands (Western Mediterranean). Fishes 2023, 8, 117. https://doi.org/10.3390/fishes8020117
Ferragut-Perello F, Ramírez-Amaro S, Tsikliras AC, Petit-Marty N, Dimarchopoulou D, Massutí E, Serrat A, Ordines F. Exploitation and Conservation Status of the Thornback Ray (Raja clavata) in the Balearic Islands (Western Mediterranean). Fishes. 2023; 8(2):117. https://doi.org/10.3390/fishes8020117
Chicago/Turabian StyleFerragut-Perello, Francesca, Sergio Ramírez-Amaro, Athanassios C. Tsikliras, Natalia Petit-Marty, Donna Dimarchopoulou, Enric Massutí, Alba Serrat, and Francesc Ordines. 2023. "Exploitation and Conservation Status of the Thornback Ray (Raja clavata) in the Balearic Islands (Western Mediterranean)" Fishes 8, no. 2: 117. https://doi.org/10.3390/fishes8020117