
Distribution of Viviparous American Fish Species in Eastern Europe on the Example of Gambusia holbrooki Girarg, 1859 and Poecilia reticulata Peters, 1859 in the Context of Global Climate Change †

 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
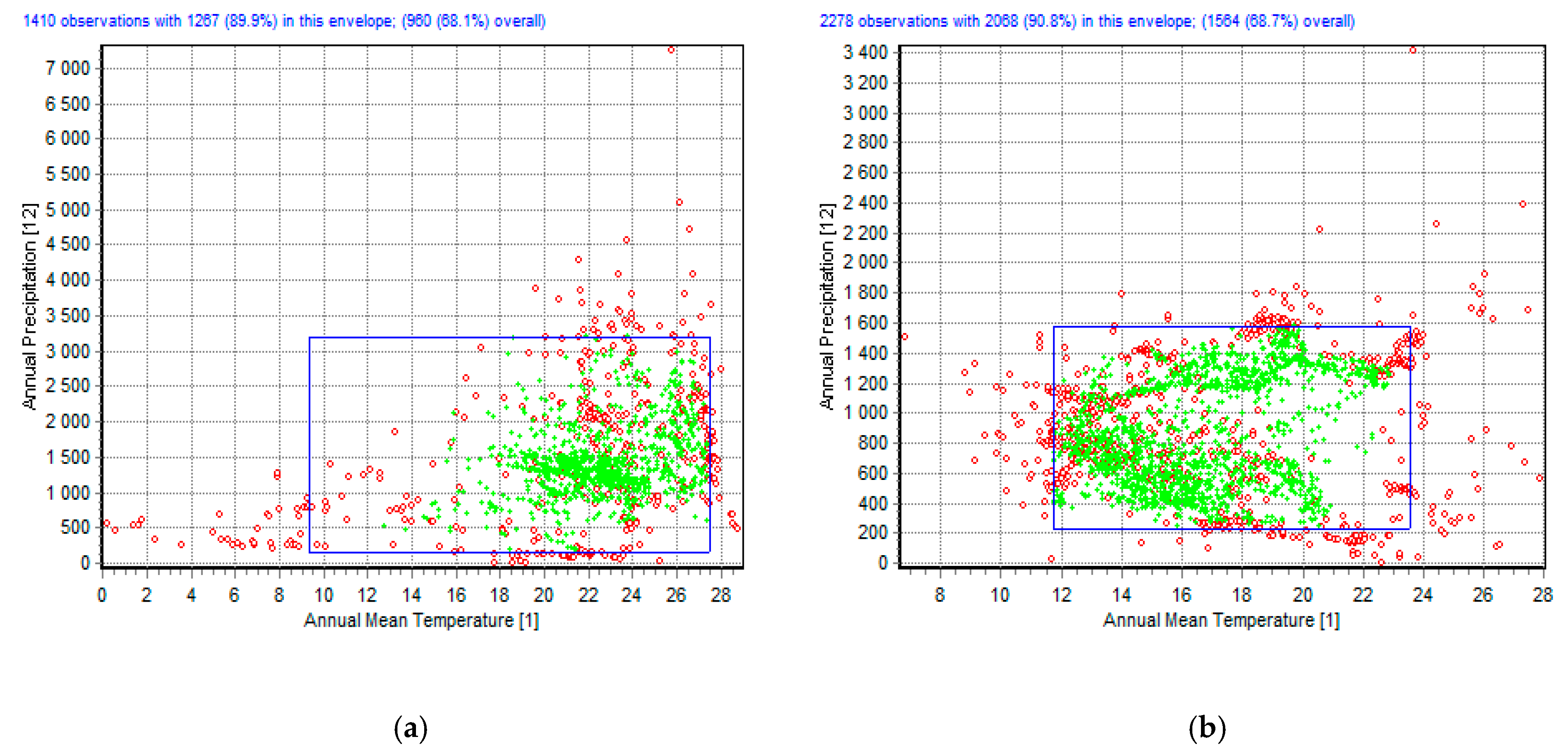
:1. Introduction
2. Materials and Methods
3. Results
3.1. Limiting Factors for P. reticulata Distribution
3.2. Limiting Factors for G. holbrooki Distribution
4. Discussion
Author Contributions
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Abbreviations
| SDM | Species distribution model |
| ROC | Receiver operating characteristic |
| AUC | Area under the ROC curve |
| CR | Critically Endangered |
| EN | Endangered |
| VU | Vulnerable |
References
- Kuybida, V.V.; Nekrasova, O.D.; Kutsokon, Y.K.; Lopatynska, V.V. Summer fish kills in the Kaniv Reservoir. Hydrobiol. J. 2019, 55, 103–106. [Google Scholar] [CrossRef]
- Pupina, A.; Pupins, M.; Nekrasova, O.; Tytar, V.; Kozynenko, I.; Marushchak, O. Species distribution modelling: Bombina bombina (Linnaeus, 1761) and its important invasive threat Perccottus glenii (Dybowski, 1877) in Latvia under global climate change. J. Environ. Res. Eng. Manag. 2018, 74, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Krumholz, L.A. Reproduction in the western mosquitofish, Gambusia affinis affinis (Baird & Girard), and its use in mosquito control. Ecol. Monogr. 1948, 18, 1–43. [Google Scholar]
- García-Berthou, E.; Alcaraz, C.; Pou-Rovira, Q.; Zamora, L.; Coenders, G.; Feo, C. Introduction pathways and establishment rates of invasive aquatic species in Europe. Can. J. Fish. Aquat. Sci. 2005, 62, 453–463. [Google Scholar] [CrossRef]
- Vidal, O.; García-Berthou, E.; Tedesco, P.A.; Garcia-Marin, J.-L. Origin and genetic diversity of mosquitofish (Gambusia holbrooki) introduced to Europe. Biol. Invasions 2010, 12, 841–851. [Google Scholar] [CrossRef]
- Kats, L.B.; Ferrer, R.P. Alien predators and amphibian declines: Review of two decades of science and the transition to conservation. Divers. Distrib. 2003, 9, 99–110. [Google Scholar] [CrossRef]
- Alcaraz, C.; Bisazza, A.; García-Berthou, E. Salinity mediates the competitive interactions between invasive mosquitofish and an endangered fish. Oecologia 2008, 155, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Padilla, D.K.; Williams, S.L. Beyond ballast water: Aquarium and ornamental trades as sources of invasive species in aquatic ecosystems. Front. Ecol. Environ. 2004, 2, 131–138. [Google Scholar] [CrossRef]
- Strecker, A.L.; Campbell, P.M.; Olden, J.D. The aquarium trade as an invasion pathway in the Pacific Northwest. Fisheries 2011, 36, 74–85. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.P.; Magurran, A.E. Multiple benefits of multiple mating in guppies. Proc. Natl. Acad. Sci. USA 2000, 97, 10074–10076. [Google Scholar] [CrossRef] [Green Version]
- Sokolov, L.I.; Sokolova, E.L.; Pegasov, V.A. Ichthyofauna of Moskva river within the city of Moscow and some data on its condition. Quest. Ichthyol. 1994, 34, 634–641. (In Russian) [Google Scholar]
- Reshetnikov, Y.S.; Bogutskaya, N.G.; Vasiljeva, E.D. List of fish-like animals and fish of fresh water of Russia. Quest. Ichthyol. 1997, 37, 723–771. (In Russian) [Google Scholar]
- Slynko, Y.V.; Kyiashko, V.N.; Yakovlev, V.N. List of Fish-Like Animals and Fish of Volga River Basin. In Catalogue of Plants and Animals of Volga River Basin; ИБBB PAH: Yaroslavl, Russia, 2000; pp. 252–277. (In Russian) [Google Scholar]
- Yakovlev, V.N.; Slynko, Y.V.; Kyiashko, V.N. Annotated list of Cyclostomata and Fish of water bodies of Upper Volga. In Ecological problems of Upper Volga; ИБBB PAH: Yaroslavl, Russia, 2001; pp. 52–69. (In Russian) [Google Scholar]
- Kozynenko, I.I.; Tytar, V.M. Bioclimatic modeling of the European distribution of the invasive Asian tiger mosquito, Aedes (Stegomyia) albopictus (Skuse, 1895), with special reference to Ukraine. Rep. Nat. Acad. Sc. Ukr. 2020, 3, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Kutsokon, Y.; Nekrasova, O.; Shkamerda, V.; Loparev, S. The spread of guppy (Poecilia reticulata Peters, 1859) in the Bortnychi aeration station channel of Kyiv City. In Biodiversity Dynamics 2012: In the Abstract of Scientific Materials; Zagorodniuk, I., Ed.; DZ—Taras Shevchenko LNU: Lugansk, Ukraine, 2012; pp. 94–95. (In Russian) [Google Scholar]
- Kvach, Y.; Kutsokon, Y. The non-indigenous fishes in the fauna of Ukraine: A potentia ad actum. Bioinvasions Record 2017, 6, 269–279. [Google Scholar] [CrossRef]
- Gambusia holbrooki Girard, 1859 in GBIF Secretariat (2021). GBIF.org (2 April 2021) GBIF Occurrence Download. Available online: https://doi.org/10.15468/dl.d5e6w9 (accessed on 2 April 2021).
- Poecilia reticulata Peters, 1859 in GBIF Secretariat (2021). GBIF.org (16 June 2021) GBIF Occurrence Download. Available online: https://doi.org/10.15468/dl.tuwazq (accessed on 16 June 2021).
- Osorio-Olvera, L.; Lira-Noriega, A.; Soberón, J.; Peterson, A.T.; Falconi, M.; Contreras-Díaz, R.G. ntbox: An r package with graphical user interface for modelling and evaluating multidimensional ecological niches. Methods Ecol. Evol. 2020, 11, 1199–1206. [Google Scholar] [CrossRef]
- Phillips, S.J. A brief tutorial on Maxent. AT&T Res. 2005, 190, 231–259. [Google Scholar]
- Kriticos, D.J.; Jarošik, V.; Ota, N. Extending the suite of Bioclim variables: A proposed registry system and case study using principal components analysis. Methods Ecol. Evol. 2014. [Google Scholar] [CrossRef]
- CliMond. Available online: https://www.climond.org/ (accessed on 27 December 2020).
- Nekrasova, O.D.; Tytar, V.M.; Kuybida, V.V. GIS Modeling of Climate Change Vulnerability of Amphibians and Reptiles in Ukraine; NAS of Ukraine, Shmalgausen Institute of Zoology NAS: Kyiv, Ukraine, 2019; 204p, ISBN 978-966-02-8956-7. (In Ukrainian) [Google Scholar]
- National Geographic Russia. Available online: https://nat-geo.ru/nature/v-sochi-ozhidayut-nashestviya-malyarijnyh-komaroviz-za-massovoj-gibeli-ryby-gambuzii/ (accessed on 11 June 2020).
- Global Invasive Species Database: Gambusia Holbrooki. Available online: http://issg.org/database/species/ecology.asp?si=617&fr=1&sts=sss&lang=EN (accessed on 11 June 2020).



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nekrasova, O.; Tytar, V.; Pupins, M.; Čeirāns, A.; Marushchak, O.; Skute, A. Distribution of Viviparous American Fish Species in Eastern Europe on the Example of Gambusia holbrooki Girarg, 1859 and Poecilia reticulata Peters, 1859 in the Context of Global Climate Change. Biol. Life Sci. Forum 2021, 2, 9. https://doi.org/10.3390/BDEE2021-09398
Nekrasova O, Tytar V, Pupins M, Čeirāns A, Marushchak O, Skute A. Distribution of Viviparous American Fish Species in Eastern Europe on the Example of Gambusia holbrooki Girarg, 1859 and Poecilia reticulata Peters, 1859 in the Context of Global Climate Change. Biology and Life Sciences Forum. 2021; 2(1):9. https://doi.org/10.3390/BDEE2021-09398
Chicago/Turabian StyleNekrasova, Oksana, Volodymyr Tytar, Mihails Pupins, Andris Čeirāns, Oleksii Marushchak, and Arturs Skute. 2021. "Distribution of Viviparous American Fish Species in Eastern Europe on the Example of Gambusia holbrooki Girarg, 1859 and Poecilia reticulata Peters, 1859 in the Context of Global Climate Change" Biology and Life Sciences Forum 2, no. 1: 9. https://doi.org/10.3390/BDEE2021-09398